
3. Строение представителей Типа Кишечнополостные
Представители Типа Кишечнополостные — это многоклеточные животные, имеющие лучевую (радиальную) симметрию.
Их тело состоит из двух слоёв клеток — наружного (эктодермы) и внутреннего (энтодермы), между которыми расположена мезоглея.
В основном, кишечнополостные — хищники. Они имеют кишечную полость, где переваривается пища. Полость сообщается с окружающей средой через рот. Других отверстий нет (непереваренные остатки выбрасываются наружу через рот).
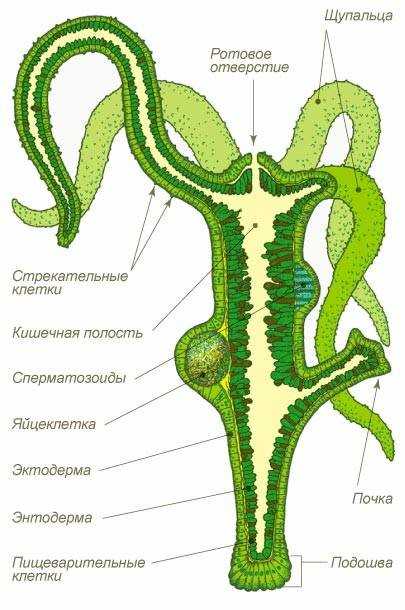
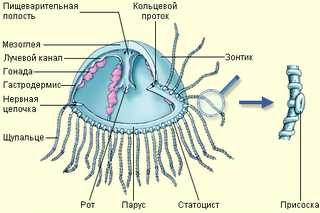
Схема строения кишечнополостных (на примере гидры пресноводной)
Обрати внимание!
Эктодерма образована эпителиально-мускульными, стрекательными, нервными, половыми и промежуточными (неспециализированными) клетками.
Энтодерма представлена пищеварительно-мускульными и железистыми клетками.
Функции клеток
1. Эпителиально-мускульные (кожно-мускульные) клетки выполнят покровную функцию, а также имеют мышечные отростки, которые обеспечивают движение кишечнополостного.
2. Стрекательные клетки имеют капсулу, заполненную ядом, который парализует жертву (нейропаралитического действия). В капсулу погружена стрекательная нить. На поверхности клетки расположен чувствительный волосок. При прикосновении к этому волоску стрекательная нить выбрасывается наружу и входит в тело жертвы.
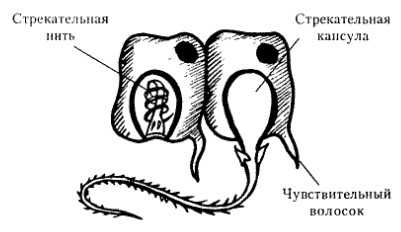
Схема строения стрекательной клетки
3. Нервные клетки имеют длинные отростки, которые вместе образуют нервную сеть. Такая нервная система называется диффузной.
Нервная система и восприятие гидрой раздражения
4. Половые клетки обеспечивают половое размножение кишечнополостных.
5. Железистые клетки продуцируют ферменты, которые переваривают пищу в кишечной полости (это внутриполостное пищеварение).
6. Пищеварительно-мускульные клетки имеют жгутики и ложноножки. Жгутики перемещают воду с частичками пищи, а образующиеся ложноножки захватывают её. Дальнейшее пищеварение происходит в пищеварительных вакуолях (это внутриклеточное пищеварение).
7. Неспециализированные (промежуточные) клетки способны превращаться в любой тип клеток и обеспечивают регенерацию (восстановление утраченных частей) кишечнополостных.
Книдоциль — чувствительный волосок стрекательной клетки кишечнополостных.
Ферменты — биологически активные вещества, которые ускоряют процессы, проходящие в клетке. Пищеварительные ферменты ускоряют процессы пищеварения.
Размножение кишечнополостных происходит половым и бесполым путями.
Бесполое размножение происходит почкованием.
В случае полового размножения из оплодотворённой яйцеклетки развивается новый организм в личиночной стадии. Прикрепившись ко дну, личинка превращается в полип. Полипы либо образовывают колонии, либо отпочковывают свободноживущих медуз. Здесь мы можем говорить о чередовании поколений: прикреплённого полипа и свободноживущей медузы.
Значение кишечнополостных
Представители Кишечнополостных — коралловые полипы образуют рифы, а иногда и целые острова — атоллы — которые представляют особые экосистемы.

Коралловый риф
Из отмерших кораллов образуются известковые горные породы. Их используют в строительстве, при декоративном оформлении жилищ.
Некоторые кишечнополостные поселяются на подвижных животных (актиния и рак отшельник). Рак перемещает актинию, а актиния защищает рака своими стрекательными клетками.
В восточных странах кишечнополостные (медузы) используются в пищу.
Яд некоторых медуз может быть опасным для человека, вызывает ожоги, а в тяжелых случаях, поражая дыхательную систему, приводит к смерти.
Источники:
Биология. Животные. 7 кл.: учеб. для общеобразоват. учреждений / В. В. Латюшин, В. А. Шапкин. — 10-е изд., стереотип. М.: Дрофа, 2009. — 302, [2] с.: ил.
Вахрушев А. А., Бурский О. В., Раутиан А. С. Биология (От амёбы до человека). 7 класс. Учебник общеобразовательной школы. — М: Баласс, 2013. — 320 с., ил. (Образовательная система «Школа 2100»).
Иллюстации:
http://drukarniacd.home.pl/flash/radial-symmetry-cnidaria
http://toazhotel.com/zhivye-obraza-zashhity.html
www.yaklass.ru
Строение представителей Кишечнополостных — Bio-learn.com
Кишечнополостные (Coelenterata) или стрекающие (Cnidaria) – богатый видами тип примитивных многоклеточных животных с радиальной симметрией тела, часто придающей им сходство с цветком. Кишечнополостные – это первые животные, у которых появились настоящие ткани.
Эти животные представляют, по сути, мешок, стенка которого состоит из наружного (эктодермы) и внутреннего (эндодермы) слоёв. Эктодерма состоит из сократительных эпителиально-мускульных и стрекательных клеток, а эндодерма – из пищеварительных эпителиально-мускульных и железистых клеток. Стрекательные клетки служат для нападения и защиты. Слои разделены мезоглеей – бесструктурным желеобразным веществом, в котором также могут находиться клетки. Дыхательная, кровеносная и выделительная системы отсутствует.
Внутреннюю часть тела занимает кишечная полость, нередко разделённая на отдельные камеры. Она открывается наружу ротовым отверстием, через которое происходит и заглатывание пищи, и экскреция. Рот обычно окружён щупальцами, служащими для захвата пищи. Пищеварение происходит как в кишечной полости, так и клетках эндодермы, захватывающих частицы пищи с помощью ложноножек.
Их тело состоит из двух слоев клеток — наружного (эктодермы) и внутреннего (энтодермы), между которыми расположена мезоглея.
В основном, кишечнополостные — хищники. Они имеют кишечную полость, где переваривается пища. Полость сообщается с окружающей средой через рот. Других отверстий нет (непереваренные остатки выбрасываются наружу через рот).
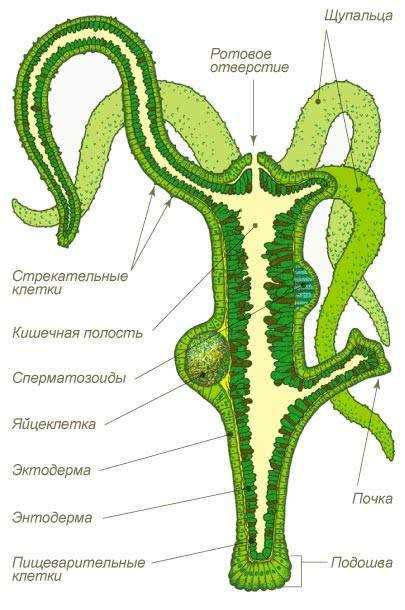
Схема строения кишечнополостных (на примере гидры пресноводной)
Энтодерма представлена пищеварительно-мускульными и железистыми клетками.
Функции клеток
1. Эпителиально-мускульные (кожно-мускульные) клетки выполнят покровную функцию, а также имеют мышечные отростки, которые обеспечивают движение кишечнополостного.
2. Стрекательные клетки имеют капсулу, заполненную ядом, который парализует жертву (нейропаралитического действия). В капсулу погружена стрекательная нить. На поверхности клетки расположен чувствительный волосок. При прикосновении к этому волоску стрекательная нить выбрасывается наружу и входит в тело жертвы.
3. Нервные клетки имеют длинные отростки, которые вместе образуют нервную сеть. Такая нервная система называется диффузной.
4. Половые клетки обеспечивают половое размножение кишечнополостных.
5. Железистые клетки продуцируют ферменты, которые переваривают пищу в кишечной полости (это внутриполостное пищеварение).
6.Пищеварительно-мускульные клетки имеют жгутики и ложноножки. Жгутики перемещают воду с частичками пищи, а образующиеся ложноножки захватывают её. Дальнейшее пищеварение происходит в пищеварительных вакуолях (это внутриклеточное пищеварение).
7. Неспециализированные (промежуточные) клетки способны превращаться в любой тип клеток, и обеспечивают регенерацию (восстановление утраченных частей) кишечнополостных.
Книдоциль — чувствительный волосок стрекательной клетки кишечнополостных.
Ферменты — биологически активные вещества, которые ускоряют процессы, проходящие в клетке. Пищеварительные ферменты ускоряют процессы пищеварения.
Размножение
Размножение кишечнополостных происходит половым и бесполым путями. Бесполое размножение происходит почкованием.
В случае полового размножения из оплодотворённой яйцеклетки развивается личиночная стадия. Прикрепившись ко дну, личинка превращается в полип. Полипы либо образовывают колонии, либо отпочковывают свободноживущих медуз. Здесь мы можем говорить о чередовании поколений: прикрепленного полипа и свободноживущей медузы.
Значение кишечнополостных
Представители Кишечнополостных — коралловые полипы образуют рифы, а иногда и целые острова — атоллы — которые представляют особые экосистемы.
Из отмерших кораллов образуются известковые горные породы. Их используют в строительстве, декоративном оформлении жилищ.
Некоторые кишечнополостные поселяются на подвижных животных (актиния и рак отшельник). Рак перемещает актинию, а актиния защищает рака своими стрекательными клетками.
В восточных странах кишечнополостные (медузы) используются в пищу. Яд некоторых медуз может быть опасным для человека, вызывая ожоги, а в тяжелых случаях, поражая дыхательную систему, приводя к смерти.
bio-learn.com
Стрекательные клетки кишечнополостных и действие их яда.
Стрекательное оружие медуз, коралов и полипов
Характернейшая особенность всех кишечнополостных, в том числе медуз, полипов, кораллов, — наличие стрекательных, или крапивных, клеток, которые развиваются из недифференцированных промежуточных клеток.
Каждая стрекательная клетка содержит хитиновую капсулу овальной или продолговатой формы. Стенки капсулы двухслойны. За счет первого (наружного) слоя на переднем конце капсулы формируется маленькая крышечка, второй же слой впячивается внутрь и дает начало тонкой спирально закрученной трубочке, называемой стрекательной нитью.
Полость капсулы заполнена ядовитой жидкостью.
На внешней поверхности стрекательной клетки имеется чувствительный волосок — книдоциль. Он представляет собой жгутик, сходный по устройству со жгутиком простейших, но, в отличие от них, неподвижный. Книдоциль окружен пальцевидными выростами цитоплазмы — микроворсинками, которые можно увидеть только с помощью микроскопа.
Малейшее прикосновение к книдоцилю вызывает возбуждение стрекательной клетки, проявляющееся в стремительном, подобном выстрелу, выворачивании стрекательной нити. В зависимости от деталей строения и способа воздействия на врага или жертву, различают множество типов строения стрекательных капсул.
Одни имеют длинную, снабженную шипами нить. При «выстреле» такой капсулы нить вонзается в тело жертвы и изливает содержимое капсулы, вызывающее местное или общее отравление.
У других капсул нить короткая, лишенная шипов. Такие нити только опутывают жертву.
Наконец, имеются липкие нити, которые удерживают добычу приклеиванием. Они же могут служить и для временного приклеивания животного при передвижении.
Стрекательные клетки располагаются по всей поверхности тела кишечнополостных, но их наибольшие скопления наблюдаются на щупальцах и вокруг ротового отверстия, т.е. в местах, где они нужнее всего.
После «выстрела» стрекательная клетка погибает и на ее месте или поблизости от него развивается новая.
Интересно, что стрекательные капсулы могут действовать и после гибели животного. Так, прикосновение к мертвым, выброшенным на берег медузам может вызвать сильные покраснения кожи, сопровождающиеся жжением.
При поедании кишечнополостных некоторыми турбелляриями и заднежаберными моллюсками, часть стрекательных клеток переходит в тело хищника. Здесь стрекательные клетки располагаются в наружных покровах и могут нормально функционировать.
Стрекательные клетки, особенно содержащие ядовитую жидкость в капсулах, — грозное оружие, которое используется кишечнополостными (медузами, полипами, кораллами) и для защиты и для нападения. Мелкие животные, соприкасаясь с кишечнополостными, приклеиваются, опутываются и пронзаются стрекательными нитями, а в их тело попадает яд, вызывающий паралич или смерть.
После этого жертва переносится щупальцами к ротовому отверстию и проглатывается. Даже довольно крупные животные часто оказываются пораженными ядом стрекательных капсул, вызывающим сильнейшую боль.
Сила воздействия яда различных видов кишечнополостных на человека неодинакова: некоторые из них совершенно безвредны, другие представляют собой серьезную опасность.
После прикосновения к нашим черноморским медузам корнеротам или аурелиям, вы почувствуете жжение, которое тем сильнее, чем нежнее кожа в месте контакта с медузой и похоже на ожог крапивой. Эти неприятные ощущения быстро проходят, не оставляя следа.
Однако в море имеется много гораздо более опасных кишечнополостных, прикосновение к которым может вызвать тяжелое заболевание и даже смерть человека. Так, многие актинии, обитающие, в том числе, и в Японском море, вызывают не только «ожоги», но и длительное опухание места контакта.
Тем не менее, наиболее неприятные последствия заключаются не в «ожогах» и опухолях, а в общем отравлении организма ядом стрекательных клеток.
У моряков, плавающих в тропических морях, уже издавна пользуется дурной славой красивая сифонофора физалия (Physalia physalis). Физалия снабжена большим, до 20 см длиной, плавательным пузырем, поднимающимся над поверхностью воды, от которого свешиваются вниз длиннейшие (до 30 м) ловчие щупальца, снабженные многочисленными стрекательными клетками.
Физалия очень ярко окрашена — пузырь отливает голубым, фиолетовым и пурпурным цветом, все свешивающиеся вниз придатки ультрамариновые. За яркую окраску физалия получила также название «португальского военного кораблика» — в средние века португальцы любили пестро окрашивать свои военные суда.
Плавающая на поверхности моря физалия видна издалека, и пловцы всегда остерегаются соприкосновения с ней, так как можно получить сильнейший «ожог», вызывающий жгучую боль. Пораженный физалией человек, даже если он отличный пловец, с трудом удерживается на воде. Вскоре после этого может наступить общее тяжелое заболевание с повышением температуры, длящееся несколько дней.
Физалия распространена исключительно в тропических морях и очень редко заносится течением и ветром в более теплые моря умеренных широт.
Нередко появляются сообщения о серьезных и даже смертельных отравлениях, вызываемых крупными сцифоидными медузами Chirodropus, Chirqsalmus, и, в особенности, Chironex из отряда кубомедуз, которые считаются одними из самых опасных существ, обитающих на нашей планете. Это тоже обитатели тропических морей. Их еще называют «морскими осами».
Высота колокола этих медуз достигает 10-15 см, по краю его сидят четыре разветвленных щупальца. Яд, содержащийся в их стрекательных клетках, попадая на кожу, вызывает омертвение эпидермиса и плохо заживающие язвы, после которых остаются глубокие рубцы.
Но самая большая опасность таится в воздействии яда на нервную систему, которое может привести к поражению дыхательного центра и вызвать смерть человека.
Впервые на ядовитость этих медуз обратили внимание в годы Второй Мировой войны в Австралии, куда было эвакуировано значительное число европейцев.
Стало известно несколько случаев загадочной гибели людей во время купания, причем на теле погибших не находили явных признаков повреждений. Долгое время тайна оставалась неразгаданной, но потом удалось установить, что причиной гибели является медуза хиродропус.
Получивший «ожог» человек захлебывался и тонул. Было отмечено, что среди пострадавших или погибших были, преимущественно, приезжие. Местные жители, особенно из числа коренного населения Австралии, могли купаться безбоязненно. По-видимому, у них выработался иммунитет к яду этой медузы.
В фауне наших дальневосточных морей также имеется один вид медуз, относящаяся к классу гидроидных, которая вызывает серьезные заболевания при соприкосновении с ней. Местные жители называют эту медузу «крестовичком» (научное название — Gonionemus vertens) за крестообразное расположение четырех темных радиальных каналов, вдоль которых тянутся четыре также темноокрашенные половые железы.
Зонтик медузы прозрачный, слабого желтовато-зеленого цвета, у отдельных экземпляров достигает 25 мм в диаметре, но обычно — 15-18 мм. На краю зонтика крестовичка имеется до 80 щупалец, способных сильно втягиваться и сокращаться. Щупальца этих медуз густо усажены стрекательными клетками, которые располагаются поясками. Посередине длины щупальца имеется присоска, при помощи которой медуза может прикрепляться к различным предметам под водой.
Крестовички обитают в Японском море и вблизи Курильских островов. Они обычно держатся на мелководье, в зарослях морской травы зостеры. В дождливые годы, когда вода у берегов сильно опресняется, медуз-крестовичков почти нет, они погибают. В засушливое лето они могут появляться массами.
Когда одно из щупалец крестовичка случайно коснется плавающего человека, медуза устремляется в этом направлении, пытаясь прикрепиться к нему присоской на щупальце. В этот момент купальщик получает сильнейший «ожог», через несколько минут кожа на теле «ожога» краснеет, покрывается волдырями. Уже через 10-30 минут наступает общая слабость, появляются боли в пояснице, затрудняется дыхание, немеют руки и ноги.
Очень сильный яд вырабатывают стрекательные клетки зоантарий — относительно мало изученных тропических шестилучевых кораллов. Отравление ядом (названным палитоксином) сказывается на деятельности сердца и всей системы кровообращения позвоночных, в том числе и человека. Яд зоантарий оказался в 100 раз сильнее яда кобры.
Статьи по теме: medusy.ru
12.1.1.2. Отравляющие и высокотоксичные вещества паралитического действия
12.1.1*2.1. Пресинаптические блокаторы высвобождения ацетилхолина
Ботулотоксин
Ботулотоксин — белок, продуцируемый микроорганизмами Clostridium botulinum. Эти бактерии способны размножаться в белковой среде в анаэробных условиях, и продуцируемый ими экзотоксин порой является причиной массовых отравлений, при использовании в пищу испорченных консервов, копченостей, грибов и т. д. (ботулизм). Впервые случай ботулизма был зарегистрирован в 1735 г. Первое описание вспышки массового отравления, обусловленного потреблением контаминированной кровяной колбасы, было сделано в Германии в 1793 г. Название — ботулизм, происходит от латинского слова botulus — колбаса (термин впервые использован для обозначения заболевания в XIX в.). В конце XIX в. Ван Эрменген связал развитие ботулизма с действием водорастворимого токсина, вырабатываемого анаэробной бактерией, названной тогда Bacillus botulinus.
Очищенный препарат экзотоксина Clostridium botulinum — ботулотоксин изучался военными специалистами США в качестве возможного отравляющего вещества (шифр — XR).
Физико-химические свойства. Токсичность
В настоящее время известны более 7 серологических типов токсина: А, В, С, D, Е, F и т. д., близких по структуре и токсической активности. Ботулотоксин представляет собой протеины с молекулярной массой 150 ООО дальтон, состоящие из двух субъединиц (MB 100 ООО и 50 ООО), соединенных дисульфидными связями. Токсин выделен в кристаллической форме. В водных растворах частично гидролизуется; устойчив к кипячению в течение часа.
Вещество проникает в организм через желудочно-кишечный тракт с зараженной водой и пищей, а при применении его в виде аэрозоля — через органы дыхания и раневые поверхности. Смертельная доза токсина для человека при алиментарном способе воздействия составляет около 50 нг/кг массы. При применении в форме аэрозоля среднесмертельная токсодоза (LQ50) — 2′ Ю-5—5 • Ю-5 г -мин/м3. Наибольшей токсичностью ботулотоксин обладает при попадании в организм через раневые поверхности (LD50 менее 1 нг/кг).
Токсикокинетика
В пищеварительном тракте ботулотоксин не разрушается протеолити-ческими ферментами и всасывается через слизистые оболочки желудка и кишечника. При ингаляции аэрозоля вещество проникает в дыхательные пути и адсорбируется на поверхности слизистой оболочки бронхов, бронхиол и альвеолоцитов, где также происходит его всасывание. Часть адсорбированного токсина мерцательным эпителием дыхательных путей выносится в ротовую полость, откуда он поступает в желудочно-кишечный тракт. Поскольку молекулярная масса токсина велика, скорость резорбции мала. Механизмы проникновения этого белкового токсина через неповрежденные слизистые оболочки не выяснены.
Циркулирующий в крови токсин постепенно разрушается протеазами плазмы. Точное время нахождения молекулы токсина в крови не известно.
При исследовании радиоизотопным методом распределения токсиканта в организме установлено, что он избирательно захватывается нервными терминалиями холинергических волокон; часть введенного токсина путем ретроградного аксонального тока транспортируется в тела нервных клеток. Об этом свидетельствует, в частности, высокое содержание 125J в телах мотонейронов спинного мозга экспериментальных животных после введения им яда, меченного этим изотопом.
Проявления интоксикации
Скрытый период интоксикации составляет от нескольких часов до суток и более (чаще до 36 ч). Продолжительность периода зависит от пути поступления токсина в организм и подействовавшей дозы. Наименее продолжителен скрытый период при попадании вещества на раневые поверхности. В клинической картине поражения выделяют общетоксический, гастроинтестинальный и паралитический синдромы. Первые симптомы — это вегетативные реакции (тошнота, рвота, слюнотечение) и признаки общего недомогания (головная боль, головокружение). Через 1—2 сут постепенно развивается неврологическая симптоматика. Усиливается слабость, появляется сухость во рту и сухость кожных покровов. Нарушается зрение (затруднена аккомодация, расширяются зрачки, выявляется их слабая реакция на свет). Основным проявлением интоксикации является постепенно развивающийся паралич поперечно-полосатой мускулатуры. Процесс начинается с глазодвигательной группы мышц (диплопия, нистагм). Ранним признаком отравления является птоз век. Позже присоединяется паралич мышц глотки, пищевода (нарушение глотания), гортани (осиплость голоса, афония), мягкого неба (речь с носовым оттенком, при попытке глотания жидкость выливается через нос). Затем присоединяется парез (а позже и паралич) мимической мускулатуры, жевательных мышц, мышц шеи, верхних конечностей и т. д. Мышечная слабость нарастает в нисходящем направлении и порой первоначально более выражена в проксимальных мышечных группах конечностей (важный диагностический признак). Токсический процесс постепенно нарастает. Иногда лишь на 10-е сут и в более поздние сроки может наступить смерть от паралича дыхательной мускулатуры и асфиксии (при тяжелых поражениях на 3—5-й день заболевания). Расстройств чувствительности при поражении ботулотоксином не бывает. Сознание у пострадавшего полностью сохранено весь период интоксикации. Нередко присоединяются острые пневмонии, токсический миокардит, сепсис (при
раневом процессе). Летальность при отравлении ботулотоксином составляет от 15 до 30%, а при несвоевременном оказании помощи может достигать 90%. По данным литературы (И. В. Маркова и соавт., 1999), кардинальными признаками ботулизма являются:
отсутствие лихорадки;
полностью сохраненное сознание;
нормальная или замедленная частота пульса;
отсутствие нарушений чувствительности;
симметричность неврологических нарушений.
Механизм токсического действия
Ботулотоксин оказывает повреждающее действие на различные структурно-анатомические образования периферической нервной системы: нервно-мышечный синапс, нервные окончания преганглионарных нейронов и парасимпатических постганглионарных нейронов. Токсины избирательно блокируют высвобождение ацетилхолина в этих структурах. Наиболее уязвимыми являются нервно-мышечные синапсы.
В опытах in vitro и in vivo, выполненных за рубежом (Пирс и соавт., 1997), установлено, что действие ботулотоксина приводит к угнетению как спонтанного, так и вызванного возбуждением нервного волокна выброса нейромедиатора в нервных окончаниях. Чувствительность постсинаптических рецепторов к ацетилхолину не изменяется. Блокада передачи сигнала не сопровождается изменением характеристик процессов синтеза и хранения ацетилхолина. При исследованиях in vitro установлено, что после добавления в инкубационную среду токсина в концентрации Ю-8 М спонтанная и вызванная активность концевой пластинки мотонейрона снижается на 90% в течение 60-90 мин. По расчетам, для блокады одного синапса достаточно 10 молекул яда. В экспериментах также установлено, что чем выше нервная активность, тем быстрее развивается блок проведения импульса в синапсах.
Полагают, что в основе эффекта лежит нарушение токсином механизма взаимодействия синаптических везикул, в которых депонирован аце-тилхолин, с аксолеммой, — необходимый этап процесса Са2+-зависимого экзоцитоза медиатора в синаптическую щель. Электрофизиологические исследования показывают, что в отличие от нормальной реакции нервно-мышечного синапса на повышение содержания кальция в инкубационной среде (дозо-зависимый выброс нейромедиатора) увеличение концентрации экстрацеллюлярного кальция с 2 до 16 мМ не приводит к усилению выброса ацетилхолина нервным окончанием, обработанным ботулотоксином. Только совместное введение в инкубат Са2+ (4 мМ и выше) и некоторых ионофоров (4-аминопиридин, гуанидин и др.) временно усиливает выброс ацетилхолина из нервных окончаний.
Действие вещества продолжительно, до нескольких недель, и потому характер взаимодействия токсина с пресинаптическими структурами-мишенями можно рассматривать как необратимое. Полагают, что восстановЛение нормальной иннервации мышц происходит в результате формирования новых синаптических контактов.
Морфологические изменения в пораженных синапсах не выявляются методами световой и электронной микроскопии.
Молекулярный механизм действия токсина окончательно не выяснен. Доказанными являются следующие представления.
Как указывалось ранее, периоду клинических проявлений предшествует скрытый период, во время которого происходит взаимодействие токсиканта с нервными окончаниями. Выделяют четыре периода действия токсина на синапс:
связывание его с плазматической мембраной холинергических нервных окончаний;
интернализация токсина путем эндоцитоза внутрь нервного окончания;
высвобождение действующей части белковой молекулы токсина и проникновение ее в цитозоль пресинаптического окончания при участии рН-зависимой транслоказы;
проявление действующей частью токсина свойств металл-зависимых эндопротеаз и разрушение специфических белков, участвующих в процессе выделения ацетилхолина из нервного окончания.
За процесс связывания с рецептором (акцептором) нервного окончания ответственна прежде всего тяжелая субъединица сложной молекулы токсина. Аффинность окончаний различных холинергических нейронов к молекулам ботулотоксина неодинакова. Наивысшим сродством обладают окончания мотонейронов, иннервирующих произвольную мускулатуру. У различных серологических типов, ботулотоксина сродство к акцепторам неодинаково.
Интернализация связавшегося с рецепторами токсина осуществляется путем эндоцитоза. В результате токсин оказывается внутри нервного окончания заключенным в мембранные везикулы, называемые эндосомами.
После проникновения внутрь нервного волокна высокомолекулярная субъединица токсина образует пору в мембране эндосомы, которая действует как канал, через который легкая цепь проникает в цитозоль пресинаптического окончания.
Теперь, свободно передвигаясь в нервном окончании, легкая цепь токсина оказывает воздействие на субстратные белки.
Биохимические исследования показали, что нейротоксины обладают цинк-зависимой эндопептидазной активностью и энзиматически расщепляют ряд белков нервных окончаний, важных для нормального высвобождения нейротрансмиттеров. Ботулотоксины А и Е расщепляют белок SNAP-25, ботулотоксин С — НРС-1 (синтаксин), токсины D и F расщепляют изоформу VAMP-1 (синаптобревин-1), токсин В — VAMP-2 (синаптобревин-2) и т. д. Протеолитическое расщепление этих специфических белков в нервных окончаниях приводит к угнетению нормального высвобождения ацетилхолина и в конечном счете к появлению основных признаков ботулизма.
Мероприятия медицинской защиты /
Специальные санитарно-гигиенические мероприятия:
использование индивидуальных технических средств защиты (средства защиты органов дыхания) в зоне химического заражения;
участие медицинской службы в проведении химической разведки в районе расположения войск; проведение экспертизы воды и продовольствия на зараженность ОВТВ;
запрет на использование воды и продовольствия из непроверенных источников;
обучение личного состава правилам поведения на зараженной местности.
Специальные профилактические медицинские мероприятия:
• проведение санитарной обработки пораженных на передовых этапах медицинской эвакуации.
Специальные лечебные мероприятия:
применение средств патогенетической и симптоматической терапии состояний, угрожающих жизни, здоровью, дееспособности, в ходе оказания первой (само- и взаимопомощь), доврачебной и первой врачебной (элементы) помощи пострадавшим;
Медицинские средства защиты
Специфическими противоядиями ботулотоксина являются противо-ботулинические сыворотки (А, В, Е). При подозрении на поражение токсином возможно профилактическое внутримышечное введение сывороток по 1000-2000 ME каждого типа с последующим наблюдением за пострадавшим в течение 10-12 дней. Решение о назначений сывороток достаточно сложно и требует участия квалифицированного специалиста, поскольку, с одной стороны, эти лекарственные средства не всегда оказываются эффективными (иные серологические типы токсина, быстрое необратимое взаимодействие яда с нервными окончаниями), а с другой — достаточно высока вероятность осложнений, связанных с их применением (анафилаксия, сывороточная болезнь).
В эксперименте in vitro нервную передачу в синапсах, нарушенную бо-тулотоксином, временно нормализует 4-аминопиридин. Полагают, что действие вещества обусловлено его способностью облегчать транспорт ионов кальция через мембрану нервных окончаний и преодолевать тем самым блок Са2+-зависимого экзоцитоза ацетилхолина. При введении летальной дозы токсина экспериментальному животному последующее назначение 4-аминопиридина отчасти восстанавливает двигательную активность животного на 1-2 ч. К сожалению, из-за высокой токсичности и кратковременности эффекта 4-аминопиридин не может рассматриваться как эффективное противоядие. Табельные средства медицинской защиты отсутствуют.
При появлении признаков угнетения дыхания необходимо предусмотреть возможность перевода пострадавшего на искусственную вентиляцию легких.
12.1.1.2.2. Блокаторы Na+-ионных каналов возбудимых мембран
Сакситоксин. Тетродотоксин
В строгом смысле слова вещества этой группы не относятся к «чистым» нейротоксикантам, поскольку, блокируя ионные каналы, действуют на возбудимые мембраны всех типов клеток организма: нервных, мышечных, железистых. Порой не возможно решить, поражение какой из структур является ведущим в патогенезе острой интоксикации. Тем не менее внешние признаки тяжелого поражения очень напоминают действие миорелаксантов, традиционно относимых к группе нейротоксикан-тов. И поэтому, хотя механизм действия веществ иной, представляется целесообразным рассмотреть их свойства в данном разделе. Достаточно хорошо изученными представителями группы являются сакситоксин и тетродотоксин, признаки поражения которыми, по сути, одинаковы. Боевое применение токсинов маловероятно, однако эти вещества рассматривались в качестве возможных диверсионных средств (В. В. Мясников, 1989; Франке, 1973). В 60-70-х гг. XX в. свойства токсинов активно изучались военным ведомством США.
Физико-химические свойства. Токсичность
Сакситоксин. В 1957 г. Шантцем с соавт. были изучены свойства так называемого «паралитического яда моллюсков» — одного из наиболее токсичных веществ небелковой природы. По названию морского моллюска, из ткани которого токсикант выделили (Saxidomus), вещество получило название сакситоксин. Позже было установлено, что в организме животных сакситоксин не синтезируется, а поступает туда с одноклеточными (жгутиковые) вида Gonyaulax catenella, которыми моллюски питаются. Количество вырабатываемого простейшими вещества колеблется в очень широких пределах и зависит от географического региона, времени года и других условий.
В случае массового размножения Gonyaulax целый ряд моллюсков поглощают их в большом количестве и концентрируют в своих тканях токсин, который для них практически безвреден. Становясь при этом ядовитыми, моллюски, съедобные для человека в обычных условиях, при использовании в пищу, вызывают случаи массового отравления людей.
Сине-зеленые водоросли пресноводных водоемов также синтезируют сакситоксин. Наблюдались случаи отравления скота водой, зараженной этими водорослями.
Сакситоксин (МВ-372) — аморфный, хорошо растворимый в воде спирте, метаноле, ацетоне порошок. Вещество устойчиво в водных растворах. Химическое строение представлено на рис. 61. Молекулы сакси-токсина, выделенные из разных источников, не полностью идентичны

NH2
Рис. 61. Структура молекулы сакситоксина
Расчетная смертельная доза сакситоксина для человека составляет по разным данным 0,004—0,01 мг/кг. Токсичность для мышей при внутри-брюшинном способе введения — около 0,009 мг/кг (с регистрацией гибели в течение 30 мин). При назначении вещества через рот смертельная доза — 0,25 мг/кг.
Тетродотоксин обнаружен в тканях целого ряда живых существ, среди которых рыбы (более 70 видов, в том числе семейства Tetrodontidae — четы-рехзубообразные), лягушки (3 вида), моллюски (1 вид). В Японии, где представитель четырехзубообразных, рыба Фугу, является деликатесом, десятки людей ежегодно отравляются в результате неумелого приготовления блюда.
Вещество выделено в чистом виде, структура его изучена (рис. 62). Это — бесцветный порошок, хорошо растворимый в воде. Раствор стабилен при комнатной температуре. Молекулярная масса — 319,3.
о

Рис. 62. Структура молекулы тетродотоксина
бе в1еле^ия otn^m ^ 6елыхттмыи«* «Ри внугрибрюшинном спосо- v ccSJ ‘ МГ/КГМаССЬЬДозав °’005 ^Асг (подкожно) вызывает
Jcf ™ Р ^ наРУшение дыхания, а дозы более 0,006 мг/кг в течение часа приводят к гибели в результате прекращения дыхания и асфиксии
Токсикокинетика
Через неповрежденную кожу вещества не проникают. Опасность представляет попадание токсинов на раневые поверхности, а также (прежде всего) потребление воды и пищи, зараженной ядами. Вещества быстро абсорбируются в кишечнике и столь же быстро выводятся из организма с мочой. Детально токсикокинетика токсинов не изучена. Дискуссионным остается вопрос о способности веществ проникать через гематоэнцефалический барьер.
Основные проявления интоксикации
Независимо от способа поступления в организм симптомы отравления практически одинаковы.
Спустя 10—45 мин появляются тошнота, рвота, боли в животе, понос. Ранними признаками поражения являются парестезии в области рта, губ, языка, десен, распространяющиеся на область шеи, покалывание, ощущение жжения кожи конечностей. Позже развиваются бледность кожных покровов, беспокойство, общая слабость, онемение конечностей, возникает ощущение невесомости тела. Зрачок сначала сужен, затем расширяется. В тяжелых случаях взгляд фиксирован, зрачковый и корнеальный рефлексы отсутствуют, появляются признаки бульбарных нарушений: затруднение глотания, речи (иногда — афония), нарастают брадикардия и гипотензия, отмечаются гиперсаливация, профузная потливость, понижение температуры тела. Дыхание учащается, становится поверхностным, развивается цианоз губ и конечностей.
Двигательные расстройства проявляются все отчетливее: появляются подергивания отдельных групп мышц, тремор, координация движений нарушается. Начавшись в области конечностей, постепенно развивающийся паралич распространяется на другие мышечные группы, охватывая все большие группы мышц. Сознание, как правило, сохраняется весь период интоксикации. Смерть наступает от паралича дыхательной мускулатуры и асфиксии в течение 6—24 ч от начала интоксикации. Если больной выживает, в течение последующих суток наступает практически полная нормализация состояния, в большинстве случаев, без отдаленных последствий.
Помимо типичной паралитической формы выделяют также гаст-ро-интестинальный и аллергический варианты течения отравления. Первый вариант проявляется признаками общего недомогания, чувством жажды, саливацией, болями в животе, тошнотой, рвотой, поносом.
Аллергическая форма отравления развивается у отдельных лиц с повышенной чувствительностью к токсинам. Характерно появление экзантем (эритематозная форма). Иногда на коже и слизистых оболочках образуются пузыри.
Механизм токсического действия
Тетродотоксин, как и сакситоксин, оказывает избирательное действие на возбудимые мембраны нервов и мышц. Как известно, градиент концентрации ионов между внутренней и внешней средой клетки формирует потенциал покоя возбудимой мембраны, равный примерно 90 мВ (табл. 52).
Таблица 52
Распределение ионов внутри и вне возбудимых клеток, мМ/л (по Katz, 1971)
Ионы | Мышечная клетка | Нервная клетка | ||
снаружи | внутри | снаружи | внутри | |
Na+ | 120 | 9,2 | 460 | 50 |
К+ | 2,5 | 140 | 10 | 400 |
ci- | 120 | 3-4 | 540 | 40-100 |
Градиенты концентраций калия и хлора примерно уравновешивают друг друга. Поэтому проницаемость мембраны для этих ионов хотя и ограничена, но относительно высока. Проницаемость натриевых каналов в покое ничтожно мала. Более того, Na+ постоянно «выкачивается» за пределы нейрона с помощью энергозависимых механизмов против высокого электрохимического градиента. Таким образом, потенциал покоя представляет собой не что иное, как готовый к использованию источник накопленной энергии, необходимой для генерации сигнала (потенциала действия). Если возбудимая мембрана деполяризуется примерно на 15 мВ, электровозбудимые натриевые каналы открываются, проницаемость их для ионов резко возрастает, Na+устремляется в клетку, разница потенциалов по обе стороны мембраны падает, деполяризация мембраны еще более усиливается, формируется потенциал действия и возбуждение передается по нервному (или мышечному) волокну. Затем в течение около 0,8 мс потенциал на мембране возвращается к исходному уровню, главным образом за счет выхода ионов калия из клетки. Усиление проницаемости для К+ необходимо для полной реполяризации мембраны и восстановления исходного потенциала покоя. При этом восстанавливается и исходная проницаемость мембраны для натрия.
Тетродотоксин и сакситоксин полностью блокируют проникновение ионов Na+ по ионным каналам возбудимых мембран внутрь клеток. При этом становится невозможным формирование потенциала действия возбудимых мембран — нарушается проведение нервных импульсов по нейронам, сокращение миоцитов. В эксперименте показано, что вещества действуют только при экстрацеллюлярной аппликации. В соответствии с расчетами одна молекула токсинов полностью блокирует один ионный канал. Полагают, что взаимодействие токсикантов с белковыми молекулами, формирующими ионный канал, осуществляется за счет группы гуанидина, содержащейся в структуре как сакси-, так и тетродотоксина. Взаимодействие токсинов с белками ионных каналов обратимо. In vitro яды могут быть удалены с поверхности возбудимой мембраны простым отмыванием биопрепарата.
Исчерпывающих данных о причинах развивающихся эффектов нет. Так, до конца не определено, возбудимые мембраны каких структур, нервных клеток (ЦНС, периферии) или миоцитов, являются более чувствительными к действию токсинов. Так, по мнению одних исследователей,’ остановка дыхания является следствием действия токсинов на нейроны
дыхательного центра, другие полагают, что основным является нарушение проведения нервного импульса по дыхательным нервам или возбудимости дыхательных мышц. Вероятно, более справедливо последнее предположение, поскольку электровозбудимость диафрагмы блокируется меньшими дозами токсикантов, чем проведение нервного импульса по диафрагмаль-ному нерву. Кроме того, в опытах на анестезированных кошках показано, что при введении вещества в смертельной дозе проведение нервных импульсов по диафрагмальному нерву не прекращается даже тогда, когда элект-ромиограмма диафрагмальной мышцы уже безмолвствует.
Развивающееся снижение артериального давления также связывают как с блокадой проведения нервных импульсов по симпатическим нервным волокнам, так и с параличом гладкомышечных клеток сосудистой стенки. Сердечная мышца вовлекается в процесс при введении токсиканта лабораторным животным в дозе 0,007 мг/кг.
Нарушение чувствительности (парестезии с последующим онемением) — следствие поражения возбудимых мембран чувствительных нейронов.
Многие центральные эффекты, такие как атаксия, головокружение, нарушение речи и т. д., могут быть связаны с действием вещества непосредственно на нейроны ЦНС.
Мероприятия медицинской защиты
Специальные санитарно-гигиенические мероприятия:
участие медицинской службы в проведении химической разведки в районе расположения войск; проведение экспертизы воды и продовольствия на зараженность ОВТВ;
запрет на использование воды и продовольствия из непроверенных источников.
Специальные лечебные мероприятия:
• своевременное выявление пораженных;
, . • применение средств патогенетической и симптоматической терапии состояний, угрожающих жизни, здоровью, дееспособности, в ходе оказания первой (само- и взаимопомощь), доврачебной и первой врачебной (элементы) помощи пострадавшим;
• подготовка и проведение эвакуации. Медицинские средства защиты
В порядке оказания доврачебной и первой врачебной помощи у пострадавшего необходимо вызвать рвоту, провести зондовое промывание желудка. Специфических средств профилактики и терапии интоксикации нет. Поскольку при тяжелых формах поражения единственным надежным способом сохранения жизни является перевод пострадавшего на искусственную вентиляцию легких, необходимо принять меры к скорейшей эвакуации пострадавших в лечебные учреждения. В случае сохранения жизни прогноз благоприятный: выздоровление бывает быстрым и полным.
ЗОВ
3Q9
studfiles.net
10 ядов животного мира с крайне специфическим действием
Яд — крайне эффективная вещь, когда нужно справиться с существом, превышающим тебя по размеру раз эдак в сто. Именно потому к нему чаще всего прибегают маленькие существа — насекомые, паукообразные, моллюски. Люди изобрели немало противоядий, чтобы бороться с распространёнными ядами, но некоторые из них настолько специфичны, что лучше попросту избегать их любой ценой.
Специфические яды не обязательно самые смертоносные (хотя такие тоже попадаются), но эффекты от них могут быть весьма долговременны, мучительны и разнообразны в худшем смысле этого слова. Лишний повод не касаться подозрительно выглядящих пауков, лягушек и гусениц.

Сиднейский лейкопаутинный паук вырабатывает яд атракотоксин, атакующий нервную систему человека и других приматов. Любопытно, что для большинства остальных млекопитающих этот яд совершенно безвреден. Хотя от укуса сиднейского паука имеется надёжное противоядие, боль от него вызывает ощущение разрывающихся лёгких.

Parabuthus transvaalicus — скорпион, населяющий пустыни африканского континента. Он способен разбрызгивать яд на расстояние до метра, но также не прочь ужалить обидчика. Его отрава вызывает у человека судороги и спазмы, заставляя нервно дёргать конечностями на протяжении суток.

Жаба-ага, одна из крупнейших жаб мира, умеющая метко плеваться опасным психоделическим ядом. Кроме галлюцинаций, он также вызывает обильное слюноотделение, конвульсии и рвоту — далеко не самое приятное состояние.

Lonomia obliqua — бразильская гусеница бабочки из семейства павлиноглазок, вооружённая уникальным ядом. От его воздействия в организме человека возникает геморрагический синдром, что может привести к серьёзным нарушениям в работе внутренних органов и даже смерти.

Пауки-бегуны из рода Phoneutria атакуют и защищаются с помощью яда, содержащего нейротоксин PhTx3, приводящий к постепенному параличу мышц и удушью. Особую опасность придаёт тот факт, что сам укус относительно безболезнен, и человек не всегда обращает на него необходимое внимание.

Конусы — хищные морские моллюски, специализирующиеся на парализующем яде моментального действия. Задохнуться напрямую от него не получится, а вот случайно утонуть, получив дозу, вполне реально.

Древолазы знамениты как своей прекрасной яркой окраской, так и опаснейшим ядом батрахотоксином. Он напрямую атакует сердечные мышцы, вызывает аритмию и остановку сердца. Противоядия к нему не существует, хотя и отравиться таким образом довольно сложно.

Медуза ируканджи крайне коварна — она не превышает размером 25 миллиметров, а её укус практически незаметен. Действие её яда напоминает странную болезнь, отчего получило название «синдром Ируканджи». После таких «восхитительных» синдромов, как постоянная рвота, отёк лёгких и чудовищная боль по всему телу наступает либо выздоровление, либо смерть.

Двуцветный питоху — очень красивая и страшно ядовитая птичка из Новой Гвинеи, получающая батрахотоксин из поедаемых ей жуков. Питоху неопасны для человека, но в руки их брать не рекомендуется — можно заработать химический ожог.

Синекольчатый осьминог при угрозе жизни может наградить обидчика отравленным укусом, ведущим к чуть ли не моментальному параличу и смерти. Недавно выяснилось, что токсический яд вырабатывается симбиотическими бактериями, живущими в слюне моллюска.
www.popmech.ru
Отравляющие вещества нервнопаралитического действия
К ОВ нервнопаралитического действия относятся фосфорорганические вещества (ФОВ), обладающие резко выраженным действием на нервную систему человека.
Табун (химическое название диметиламид этилового эфира цианфосфиновой кислоты) представляет собой бесцветную, прозрачную жидкость со слабым фруктовым запахом. Технический табун — красно-бурая жидкость с удельным весом 1,06, температура кипения +103—104° (при 10 мм ртутного столба) и замерзания — 40°. Табун может применяться как нестойкое и как стойкое ОВ для заражения атмосферы (в виде паров и тумана) и территории в капельножидком состоянии.
Гитлеровцы предполагали применить это ОВ в авиационных бомбах, артснарядах, путем поливки с самолетов и в виде других средств химического нападения.
В воде табун растворяется плохо, медленно разлагаясь с выделением синильной кислоты. Хорошо растворяется в бензоле, ацетоне, спирте, дихлорэтане.
При действии растворов щелочей (мыльных растворов, гашеной извести, аммиачной воды, настоев золы и т. д.) табун разлагается довольно быстро. Указанные щелочные вещества могут быть использованы с успехом для дегазации предметов и местности, зараженных табуном.
Табун, как и другие ФОВ, относится к сильнодействующим на нервную систему человека и животного ядам, вызывающим в первую очередь судороги и паралич.
В связи с указанными токсикологическими свойствами ФОВ получили названия ОВ нервнопаралитического действия. К ним относится также зарин. Особенностью ФОВ является их высокая токсичность. Насколько токсичны ОВ типа табун, можно судить по следующим данным. Самым ядовитым ОВ считалась синильная кислота, смертельная концентрация которой около 0,5 мг в литре. При заражении воздуха зарином достаточно даже такой концентрации, как 0,005 мг/л, т. е. в 100 раз меньше, чтобы вызвать потерю людьми работоспособности на срок до недели и более.
Табун и зарин легко проникают в пористые материалы — дерево, одежду, кожу.
При поражениях фосфорорганическими веществами появляются судорожные сокращения мышц, которые обусловливаются расстройством функций центральной нервной системы, возникают также расстройства дыхания.
Одним из первых признаков отравления являются нарушение зрения, сужение зрачков, слезотечение, боли в области глаз. Длительное нарушение зрения, связанное с миозом и потерей аккомодации, вызывают даже такие концентрации, как 1 : 100 000 (1 л ОВ на 100 000 л воздуха). При Вдыхании воздуха, содержащего больше ОВ, появляется вскоре головная боль, головокружение, мышечная слабость, кашель с мокротой, слюноотделение, рвота. Через 10—15 минут после отравления наступают судороги, которые длятся несколько минут, затем прекращаются, но через некоторое время приступ судорог вновь возобновляется. Смерть наступает от остановки дыхания. В зависимости от дозы ОВ, поступившей в организм, смерть может наступить в первые же минуты после отравления.
При попадании ФОВ на кожу раздражения не ощущается, однако яд легко всасывается в организм и вызывает отравление. Первые признаки отравления, как правило, проявляются уже через несколько минут после попадания яда на кожу. В отличие от отравления через легкие кожнорезорбтивные отравления могут протекать и без сужения зрачков, что является характерным признаком отравления при вдыхании зараженного воздуха ОВ типа табун.
При попадании небольших количеств ФОВ в желудок с водой либо пищей признаки отравления могут проявиться через некоторое время, спустя 20—30 минут появляются рвота, понос, угнетение, вялость. В случаях тяжелого отравления появляются судороги, расширяются зрачки, нарушается дыхание. Смерть наступает от остановки дыхания и сердечной деятельности.
Зарин — жидкость бурого цвета с удельным весом 1,1, примерно в 20 раз ядовитее табуна, запаха почти не имеет, более летуч, чем табун. Температура кипения + 146°, замерзания —37°, смешивается во всех пропорциях с водой и со многими растворителями.
Зарин является судорожно-паралитическим ОВ и по токсичности приближается к самым ядовитым веществ вам.
Отравление зарином может произойти при вдыхании его паров, а также путем проникновения в капельножидком состоянии через кожные покровы и при попадании в желудочно-кишечный тракт. Первым признаком поражения является сильное сужение зрачков, приводящее к расстройству зрения, вплоть до полной его потери. При тяжелом отравлении в первые минуты наступает затруднение дыхания, рвота, появляется дрожание и судорожное сокращение отдельных мышц.
Зарин, как и табун, от действия щелочей разлагается. Поэтому дегазаторами зарина могут быть те же вещества, что используются для дегазации табуна.
Значительно токсичнее зарина заман (в 2—3 раза), представляющий собой бесцветную жидкость со слабым запахом канифоли. Температура кипения около 260° (он менее летуч, чем зарин). Пары его в 6 раз тяжелее воздуха.
По химическому составу заман относится к фосфорорганическим соединениям — эфир метилфторфосфиновой кислоты.
Окись углерода — бесцветный газ без запаха. При температуре —191° сжижается. Образуется окись углерода при неполном сгорании различных органических веществ, в том числе при взрыве фугасных авиационных бомб. Особую опасность представляет окись углерода в закрытых, плохо вентилируемых помещениях. Содержание окиси углерода, например, при взрыве фугасной авиабомбы внутри здания может доходить до 70%, что создает смертельную концентрацию.
Окись углерода при вдыхании попадает через легкие в кровь и, входя в состав гемоглобина крови, нарушает кислородный обмен, вызывая таким образом кислородное голодание. При отравлении наблюдаются шум в ушах, головокружение, одышка, тошнота, рвота, судороги, удушье, чувство слабости, потеря сознания. При несмертельных концентрациях через 1—2 суток наступает выздоровление.
Обнаруживается окись углерода в воздухе специальной индикаторной трубкой. Для защиты на противогазовую коробку дополнительно навинчивается гопкалитовый патрон (без гопкалитового патрона противогаз от окиси углерода не защищает).
wushu-zentrum.com
Яды — высокоточное оружие: компьютерное исследование природных нейротоксинов
Биологическая эволюция — общая форма существования живой материи. При детальном рассмотрении оказывается, что виды почти никогда не эволюционируют поодиночке: обычно в этом принимают участие их экологические партнеры, и изменение происходит в парах паразит—хозяин или хищник—жертва. Более того, коэволюцию часто можно проследить на молекулярном уровне, когда один вид совершенствует систему нападения, а второй вслед за ним — систему защиты (и наоборот). Параллель с холодной войной здесь настолько очевидна, что соответствующий феномен даже получил название эволюционной «гонки вооружений». Примером наиболее эффективных и элегантных систем нападения служат животные яды, содержащие в своем составе нейротоксины — вещества, воздействующие на нервную систему и мышцы жертвы.
Несмотря на детальную проработку теории эволюции профессионалами и многочисленные примеры, вошедшие в учебники и популярную литературу [1], воплощение эволюционного процесса зачастую настолько причудливо, что исследование изменений в живых организмах может быть интересно далеко не только ученым-теоретикам. Известно, что у эволюции нет воли и цели: живые организмы меняются, чтобы оставить максимальное потомство, а не чтобы достигнуть абстрактного совершенства [2]. Так, эволюционно прогрессивным признаком считается то, что позволяет лучше приспособиться к окружающим условиям в данный момент, а не красота или сложность устройства сами по себе.
Например, антарктическая ледяная рыба утратила гены гемоглобина из-за того, что условия ее обитания — нулевая или даже отрицательная (!) температура воды, растворимость кислорода в которой достаточно высокая, чтобы обеспечивать ткани этим газом просто за счет диффузии и усиления кровотока. Можно сказать, что в этом случае рыба «экономит» на гемоглобине [3]. Другие рыбы, которые живут в пещерах и никогда не выплывают на белый свет или обитают на очень больших глубинах, почти всегда за ненадобностью теряют зрение. Степень упрощения паразитических организмов вообще поражает воображение, причем в случае бактерий это приводит к кардинальному упрощению всего генетического аппарата: например, геном микоплазмы содержит всего около 500 генов, а у облигатного внутриклеточного симбионта Candidatus Carsonella ruddii и вовсе около 180 генов [4]. И всё перечисленное — прогрессивные признаки, поскольку они позволили упомянутым организмам максимально адаптироваться к обстановке, в которой они обитают. Более известный (и более «благородный») пример — эволюция человека, где развитие пошло по пути усложнения нервной системы и способности к коммуникации, а также к становлению богатой культуры, современное состояние которой мы имеем удовольствие ежедневно наблюдать вокруг. Одновременно возросла роль полового отбора и образования семей [5], — и все это тоже проделки эволюции [6].
В нашей статье речь пойдет об особой разновидности эволюционного процесса, которую можно назвать «гонкой вооружений» и в которой участвуют две стороны: нападающая и обороняющаяся (или «догоняющая» и «убегающая»). Чрезвычайно любопытно наблюдать «гонку» на молекулярном уровне. Поскольку изменение одной стороны согласовано с изменением второй, то и гонку вооружений иногда можно довольно подробно проследить по «молекулярной летописи» процессов, происходящих внутри живых организмов.
Уроки холодной войны
Вторая половина XX века, кроме замечательных открытий в молекулярной биологии, была ознаменована холодной войной — глобальной конфронтацией между социалистическим и капиталистическим строями. Это противостояние породило эффект гонки вооружений — процесс борьбы за военное превосходство с положительной обратной связью, вызвавший гипертрофию военно-промышленных комплексов СССР и США.
Похожий эффект наблюдается и в процессе коэволюции двух видов, связанных друг с другом в системе «хищник—жертва» или «паразит—хозяин»: изменение одного из видов неизбежно влечет изменение второго для сохранения паритета или получения превосходства. Хорошо известный пример эволюционной гонки вооружений — газель и гепард, каждому из которых приходится бегать из поколения в поколение все быстрее и быстрее, чтобы одному ускользнуть из пасти, а другому — остаться сытому.
Эволюционная гонка вооружений, или принцип Черной королевы
Подчеркивая параллель с холодной войной, биологи ввели понятие эволюционной гонки вооружений (англ. evolutionary arms race), приписывая этому процессу роль важнейшей движущей силы эволюции [7]. Более строго это понятие сформулировано в форме «принципа Черной Королевы», утверждающего, что в коэволюционных отношениях каждому из видов приходится «бежать со всех ног, чтобы только остаться на том же месте» (эта цитата из «Алисы в Зазеркалье» удачно передает суть принципа). Прямое экспериментальное подтверждение принципа Черной королевы получено на микроскопических рачках дафниях и их паразитических бактериях: «воскресив» из донного ила несколько поколений тех и других, биологи показали, что самые опасные для дафний паразиты — современные им, тогда как «прошлые» и «будущие» популяции бактерий заражали рачков с меньшей эффективностью [8].
Око за око, ген за ген
Хорошим примером эволюционной гонки вооружений являются растения с их паразитами. Во многих случаях наблюдается строгое соответствие системы устойчивости растения системе вирулентности его паразита. Более того, это соответствие сводится всего лишь к двум генам: гену растения, позволяющему противостоять заражению (гену резистентности, R) и гену паразита, необходимому для инфекции (гену авирулентности, Avr). Такие межвидовые отношения получили название «ген за ген» (англ. gene-for-gene) [9]. Растения, содержащие ген R, оказываются устойчивыми по отношению к паразитам с соответствующим геном Avr. Как правило, гены резистентности растений кодируют белки-рецепторы, регистрирующие появление паразита. Теперь паразит стремится изменить свой Avr-ген так, чтобы ускользнуть от узнавания рецептором растения. И наоборот, растение изменяет свой R-ген, чтобы по-прежнему детектировать заражение.
Вариация фаз
Одно из наиболее прогрессивных эволюционных приобретений высших позвоночных — развитая иммунная система, основанная на принципе комбинаторики и позволяющая противостоять практически любому чужеродному организму [10], [11]. Однако бактерии и вирусы на сдаются, им тоже есть что предъявить из своего арсенала. Хорошо известный пример — постоянно мутирующий вирус гриппа, к которому необходимо получать всё новые и новые вакцины [12]. Другой, не менее поразительный, пример — явление «смены фаз» у бактерий, заключающееся в случайном изменении фенотипа с высокой частотой, намного превышающей частоту обычных мутаций. Так, сальмонеллы используют этот прием для замены белка флагеллина, из которого построены бактериальные жгутики и который служит сигналом для запуска иммунологических реакций. Только у хозяина разовьется иммунный ответ, как сальмонелла меняет тип флагеллина и ускользает [13]!
Ядовитый арсенал
Пожалуй, самым эффективным средством как нападения, так и защиты служат яды, которые в процессе эволюции научились вырабатывать разнообразнейшие животные: многие кишечнополостные, членистоногие, моллюски, хордовые и другие (рис. 1). Присутствующие в ядах молекулы называют токсинами, а в том частном (но распространенном) случае, когда мишенью их действия служит нервная система и/или мышцы, — нейротоксинами.

Рисунок 1. Разнообразие ядовитых животных. На филогенетическом дереве многоклеточных отмечены типы животных, среди представителей которых встречаются ядовитые. Вокруг дерева расположены фотографии ядовитых животных (сверху — позвоночных, слева — членистоногих, слева внизу — иглокожих, справа вверху — моллюсков, справа внизу — губок, кишечнополостных и некоторых червей).
Состав яда животных различен: если, допустим, в яде пчелы присутствуют всего два основных компонента — мембраноактивный пептид мелиттин и гидролизующий липиды фермент фосфолипаза A2, — то в яде пауков, скорпионов, морских анемон и конусов, а также змей содержатся десятки, а иногда сотни или даже тысячи компонентов различной химической природы. Наблюдаемое в ядах разнообразие компонентов одного структурного типа сегодня принято описывать термином «эволюционно отредактированная комбинаторная библиотека» [14]. Эти молекулы обладают различной эффективностью и специфичностью в отношении разных рецепторов, а результирующая смесь токсинов эффективна в отношении широкого круга мишеней.
Эволюционное преимущество богатого арсенала — способность «следовать» за жертвой: если мишень действия основного компонента яда начнет ускользать (например, рецептор мутирует), в яде с большой вероятностью обнаружится слабо представленный, но более активный по отношению к новой форме рецептора токсин, и теперь уже ему суждено будет стать основой «вооружения» у будущих поколений ядовитых хищников.
«Всенепременнейше захватите телеграф!»
Как уже было сказано, частой мишенью действия ядов является нервная система, а поэтому основа арсенала многих ядовитых гадов — нейротоксины. Связано это, по-видимому, с тем, что нарушить работу сложной системы проще всего, уничтожив коммуникацию — что было отмечено еще в работах Ленина, рекомендовавшего в случае захвата власти в первую очередь взять под контроль телефон, телеграф и железнодорожные станции. «Телеграфом» нашего организма является нервная сеть, передача сообщений в которой основана на феномене потенциала действия, распространяющегося вдоль мембран нейронов. В состоянии покоя поддерживается определенная разница концентраций ионов (Na+, K+, Ca2+, Cl−) внутри нейронов и снаружи, и формируется потенциал покоя [15]. Рабочими элементами потенциала действия служат ионные каналы — мембранные белки, пропускающие те или иные ионы по команде. В частности, начинается потенциал действия с активации потенциал-чувствительных натриевых (Na+) каналов (ПЧНК), открывающихся в ответ на уменьшение трансмембранной разности потенциалов (деполяризацию). Интересно, что появление ПЧНК в эволюции животных тесно связано с дифференцировкой тканей и обособлением нервной системы. О них и поговорим подробнее, оставив другие каналы за рамками нашей статьи.
Каждый ПЧНК состоит из очень длинной полипептидной цепи (около двух тысяч остатков аминокислот), которая представлена четырьмя похожими повторами (рис. 2). При этом в пространстве канал составлен из пяти частей (или доменов). В образовании единственного порового домена (ПД) участвуют все повторы полипептидной цепи; в его центре формируется селективная для ионов Na+ пора. Четыре — по одному от каждого повтора — потенциал-чувствительных домена (ПЧД) расположены вокруг центрального ПД [16]. Функция ПЧД — реагировать на изменение мембранного потенциала и передавать команду на открытие поры.
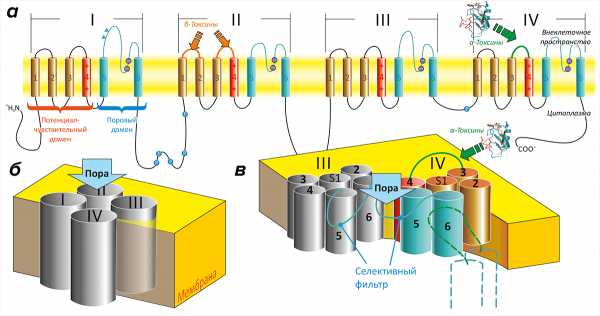
Рисунок 2. Строение потенциал-чувствительных натриевых каналов (ПЧНК). а — ПЧНК состоят из четырех похожих повторов (I–IV). Каждый повтор содержит шесть трансмембранных α-спиралей (S1—S6). Четыре спирали (S1—S4) формируют потенциал-чувствительный домен (ПЧД). Единственный поровый домен (ПД) образован восемью спиралями, по две (S5—S6) от каждого повтора. Четвертая спираль в каждом ПЧД (S4) положительно заряжена и выполняет роль сенсора потенциала. С внеклеточной стороны с ПЧД-IV и ПЧД-II связываются α- и β-токсины скорпионов. б — В пространстве повторы ПЧНК располагаются вокруг общей оси. в — Центральный ПД канала окружен ПЧД. Участок между спиралями S5 и S6 содержит «селективный фильтр», опосредующий избирательное пропускание ионов Na+.
ПЧНК являются ключевым компонентом передачи нервных импульсов, а значит и работы нервной системы вообще. Очевидно, именно этим обусловлено существование огромного числа токсинов, связывающихся с ПЧНК и так или иначе нарушающих их работу (рис. 3). В свою очередь, нарушение работы ПЧНК приводит к параличу, судорогам, а смерть наступает, как правило, от остановки дыхания. Например, тетродотоксин, содержащийся в знаменитом японском деликатесе рыбе фугу и некоторых других животных, блокирует пору ПЧНК подобно пробке. Интересно, что тетродотоксин используется чаще в целях защиты. И здесь хорошо исследован пример гонки вооружений: некоторые тритоны используют тетродотоксин для защиты от хищников, а охотящиеся на этих тритонов змеи приобретают устойчивость в результате мутации генов ПЧНК [17]. К блокаторам относятся небольшие пептиды μ-конотоксины, являющиеся оружием морских моллюсков-конусов. Местные анестетики, такие как новокаин и лидокаин, тоже являются блокаторами ПЧНК: их местное применение приводит к блокаде чувствительных нейронов.
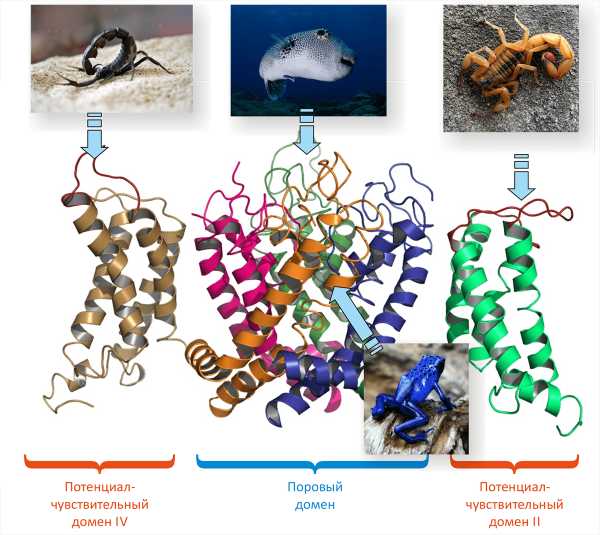
Рисунок 3. Некоторые природные нейротоксины, воздействующие на ПЧНК. Изображена модель пространственной организации канала; показан центральный ПД и два ПЧД (II и IV). Представлены фотографии ядовитых животных, служащих источником «классических» нейротоксинов. Тетродотоксин из рыбы фугу блокирует пору ПЧНК, связываясь с внеклеточной стороны. Батрахотоксин из кожи колумбийской лягушки проникает через мембрану и встраивается внутрь поры канала, вызывая его активацию. α-Токсины скорпионов связываются с ПЧД-IV и подавляют процесс естественной инактивации каналов. β-Токсины скорпионов связываются с ПЧД-II и активируют ПЧНК.
Но не все токсины блокируют канал: есть и такие, которые его активируют, то есть увеличивают время, когда канал находится в открытом состоянии. Примером таких молекул является батрахотоксин, содержащийся в секрете кожных желез некоторых Южноамериканских лягушек-листолазов. Распространенные инсектициды пиретроиды также относятся к активаторам ПЧНК.
α- и β-токсины из яда скорпионов представляют собой небольшие белки (~60—70 аминокислотных остатков), в структуре которых присутствует β-лист из трех тяжей и короткая α-спираль, скрепленные четырьмя дисульфидными связями (рис. 4). Хотя и те, и другие токсины, по сути, активируют канал, делают они это по-разному. α-Токсины мешают каналу закрыться, а β-токсины — помогают ему открыться. Соответственно и места взаимодействия с ПЧНК у этих токсинов разные: те и другие связываются с ПЧД, но для α-токсинов это ПЧД-IV, а для β-токсинов — ПЧД-II.

Рисунок 4. Модульная структура α-токсинов скорпионов. Эти токсины — небольшие белки, составленные из β-листа и α-спирали, скрепленных дисульфидными связями. Такая жесткая и эволюционно консервативная структура образует «сердцевину» молекулы, а две петли и C-конец образуют динамически подвижный «модуль специфичности» (показан пунктирным овалом), отличающийся у млеко- (слева) и инсектотоксинов (справа). Рисунок показывает результаты анализа характерных внутренних движений по данным молекулярной динамики. Цветной спектр конформаций изображает динамическую подвижность каждой из молекул.
Остановимся теперь подробнее на α-токсинах и их «взаимоотношениях» с ПЧНК. Дело в том, что в яде скорпионов присутствуют молекулы, обладающие токсическим действием по отношению к насекомым или млекопитающим (назовем соответствующие α-токсины инсектотоксинами и млекотоксинами). И те, и другие связываются с ПЧД-IV, но только у разных каналов. У насекомых это свой канал, называемый Para [18], а у млекопитающих есть целых девять разновидностей каналов, обозначаемых Nav1.1—1.9 [19]. Разные ПЧНК млекопитающих выполняют разные функции. Например, Nav1.2 характерен для центральной нервной системы, Nav1.4 — для скелетной мускулатуры, Nav1.5 — для сердца [20]. А дальше в ход идет упомянутая «комбинаторная библиотека» яда скорпионов, в которой может найтись молекула, активная по отношению к выбранной мишени. Если это происходит, «владелец» такого яда получает эволюционное преимущество по сравнению с другими, и та часть его генома, которая отвечает за состав ядовитой библиотеки, широко распространяется в будущих поколениях скорпионов.
Присутствующие в яде скорпионов нейротоксины отличаются разнообразием: некоторые из них могут быть направлены исключительно на каналы насекомых (инсектотоксины), тогда как другие действуют на каналы млекопитающих (млекотоксины). Есть также молекулы, действующие сразу и на те, и на другие ПЧНК. Что лежит в основе молекулярной эволюции этих токсинов, позволяющей им угнаться сразу за множеством ускользающих мишеней? Попробуем ответить на этот вопрос.
Компьютерный анализ выявляет «молекулярную гонку вооружений»
В Институте биоорганической химии им. академиков М.М. Шемякина и Ю.А. Овчинникова РАН было проведено исследование, направленное на выявление особенностей млеко- и инсектотоксинов из яда скорпионов, определяющих их селективное взаимодействие с соответствующими ПЧНК. Работа состояла из двух частей:
- Компьютерный анализ структуры и динамики позволил выявить характерные особенности млеко- и инсектотоксинов и предсказать активность «сиротского» токсина с неизвестной селективностью. Эта часть работы была сделана в лаборатории моделирования биомолекулярных систем.
- Затем предсказания были проверены в лаборатории нейрорецепторов и нейрорегуляторов: был синтезирован «сиротский» токсин и исследована его специфичность.
Компьютерное моделирование было основано на методе молекулярной динамики (МД) [21]. Установлено, что молекулы α-токсинов, несмотря на свой небольшой размер и жесткую структуру, состоят из двух динамических модулей. Анализ характерных движений показал, что один из этих модулей достаточно «жесткий», а другой конформационно «пластичный». Более того, движения «пластичного» модуля различаются у млеко- и инсектотоксинов (рис. 4).
Идентифицированные с помощью МД части молекул получили название «сердцевины» и «модуля специфичности». Сердцевина α-токсинов оказывается эволюционно консервативным (очень сходно устроенным у инсекто- и млекотоксинов), а модуль специфичности — вариабельным, соответствующим конкретной мишени действия. Сердцевина, таким образом, отвечает за распознавание ПЧНК «вообще», а быстро изменяющийся в эволюции модуль специфичности позволяет токсину «настраиваться» на конкретный тип канала.
Было также обнаружено, что модуль специфичности млекотоксинов существенно более гидрофильный, чем у инсектотоксинов. Эта особенность, предположительно, отражает структурные детерминанты, позволяющие токсинам селективно распознавать свои мишени. Что интересно, анализ свойств ПЧНК показал ту же тенденцию для внеклеточной части S5—S6 повтора I: у каналов млекопитающих эти области более гидрофильны, а у каналов насекомых — гидрофобны. Сопоставление результатов анализа с накопленными биохимическими данными позволило предложить любопытный характер связывания α-токсинов с ПЧНК. Сердцевинный модуль, по-видимому, взаимодействует с ПЧД-IV, в то время как модуль специфичности связывается с петлей S5—S6 повтора I. Интересно, что данные области в структуре ПЧНК сближены, что также было показано в независимых экспериментах. Идея соответствия модульной организации токсинов доменной структуре каналов имеет интересный эволюционный смысл, позволяющий нам разглядеть гонку вооружений в противостоянии ядовитых животных и их жертв. Модульная структура предположительно позволяет токсинам гибко адаптироваться к изменяющейся мишени.
Компьютерный анализ может быть использован для предсказания активности токсинов с неизученными свойствами. В частности, для токсина M9 из яда Среднеазиатского скорпиона Mesobuthus eupeus, ставшего первым α-нейротоксином из скорпионьего яда, для которого была установлена пространственная структура (кстати, это тоже было сделано в ИБХ [22]), была предсказана активность в отношении каналов как млекопитающих, так и насекомых. Биоинженерный синтез и тестирование активности этого токсина на рекомбинантных ПЧНК подтвердили высказанное предположение. Результаты представленной работы были опубликованы в журнале Journal of Biological Chemistry [23]. Эволюционное обособление модулей α-токсинов, похоже, вызвано требованиями гонки вооружений — необходимостью оперативно адаптироваться вслед за изменяющимися условиями среды: появлением новых мишеней и изменением старых. Вряд ли этот подход является универсальным, но в данном случае он позволил с новой стороны взглянуть на взаимосвязь структура—функция для биологически активных пептидов.
Биоинженерия и нейробиология
Исследования молекулярных основ эволюционной гонки вооружений имеет не только фундаментальное значение. Например, заново созданные «дизайнерские» молекулы — аналоги нейротоксинов с заданной исследователями активностью — являются идеальными инструментами исследования нервной системы. Такие молекулы позволили бы прицельно регулировать работу ионных каналов и модифицировать нервный ответ желаемым образом.
Придание нейротоксинам желаемой селективности и видоспецифичности позволит, например, создать инсектицид нового поколения. Биоинженерно изготовленный или даже внедренный в геном растений инсектотоксин позволит эффективно бороться с насекомыми-вредителями. А можно представить себе возможность создания идеально селективного инсектотоксина, действующего на вредителей и не токсичного для полезных насекомых (например, пчел).
Наконец, идеально селективные нейротоксины — уже не яды, а точно настраиваемые нейрорегуляторы — нашли бы применение в молекулярной медицине для лечения заболеваний, связанных с дисфункцией ионных каналов и называемых каналопатиями. Среди них — различные неврологические и психические расстройства, а также к патологии скелетной и сердечной мускулатуры [24].
Статья поддержана грантом РФФИ д_с-13-04-11520 и первоначально опубликована в «Науке и жизни» [25]. См. также видеозапись «ядовитого» семинара, организованного Советом молодых ученых ИБХ РАН осенью 2013 года.
Видео 1. Доклад А.А. Василевского «Что-то… про паучков», сделанный на «ядовитом» семинаре Совета молодых ученых ИБХ РАН осенью 2013 года. В этом докладе также рассказывается о непростых взаимоотношениях нейротоксинов с их молекулярными мишенями.
- Докинз Р. Самое грандиозное шоу на Земле. Доказательства эволюции. М.: «Корпус/Астрель», 2012;
- Когда б вы знали, из какого сора…;
- Возможна ли жизнь без гемоглобина?;
- S. G. E. Andersson. (2006). GENETICS: The Bacterial World Gets Smaller. Science. 314, 259-260;
- Семья — женское изобретение;
- Марков А. Эволюция человека. В 2-х томах. М.: «Династия», 2011;
- Элементы: «Гонка вооружений — двигатель эволюции»;
- Элементы: «Современные паразиты опаснее прошлых и будущих»;
- Walter Gassmann, Saikat Bhattacharjee. (2012). Effector-Triggered Immunity Signaling: From Gene-for-Gene Pathways to Protein-Protein Interaction Networks. MPMI. 25, 862-868;
- Иммунологическая Нобелевская премия (2011);
- Толл-подобные рецепторы: от революционной идеи Чарльза Джейнуэя до Нобелевской премии 2011 года;
- Гонки с вирусом: эпидемиология и экология вируса гриппа;
- M. W. van der Woude, A. J. Baumler. (2004). Phase and Antigenic Variation in Bacteria. Clinical Microbiology Reviews. 17, 581-611;
- Великому комбинатору и не снилось: комбинаторика токсинов пауков;
- Формирование мембранного потенциала покоя;
- Frank H Yu, William A Catterall. (2003). . Genome Biol. 4, 207;
- T SOONG, B VENKATESH. (2006). Adaptive evolution of tetrodotoxin resistance in animals. Trends in Genetics. 22, 621-626;
- Ke Dong. (2007). Insect sodium channels and insecticide resistance. Invert Neurosci. 7, 17-30;
- G. F. Lopreato, Y. Lu, A. Southwell, N. S. Atkinson, D. M. Hillis, et. al.. (2001). Evolution and divergence of sodium channel genes in vertebrates. Proceedings of the National Academy of Sciences. 98, 7588-7592;
- W. A. Catterall. (2005). International Union of Pharmacology. XLVII. Nomenclature and Structure-Function Relationships of Voltage-Gated Sodium Channels. Pharmacological Reviews. 57, 397-409;
- Молекулярная динамика биомолекул. Часть I. История полувековой давности;
- Vladimir S. Pashkov, Vladimir N. Maiorov, Vladimir F. Bystrov, Anh N. Hoang, Tatyana M. Volkova, Eugene V. Grishin. (1988). Solution spatial structure of ‘long’ neurotoxin M9 from the scorpion Buthus eupeus by 1H-NMR spectroscopy. Biophysical Chemistry. 31, 121-131;
- Anton O. Chugunov, Anna D. Koromyslova, Antonina A. Berkut, Steve Peigneur, Jan Tytgat, et. al.. (2013). Modular Organization of α-Toxins from Scorpion Venom Mirrors Domain Structure of Their Targets, Sodium Channels. J. Biol. Chem.. 288, 19014-19027;
- G. S.B. Andavan, R. Lemmens-Gruber. (2011). Voltage-Gated Sodium Channels: Mutations, Channelopathies and Targets. CMC. 18, 377-397;
- Чугунов А.О. и Василевский А.А. (2013). Эволюционная «гонка вооружений»: нейротоксины против ионных каналов. «Наука и жизнь». 11, 42–48;
- William A. Catterall. (2010). Ion Channel Voltage Sensors: Structure, Function, and Pathophysiology. Neuron. 67, 915-928;
- О чем не знал Гальвани: пространственная структура натриевого канала;
- Калиевый канал in silico.
biomolecula.ru