
Иж 27 общая длина. Иж 27м: технические характеристики
Содержание
- 1 ИЖ-27 – самая популярная советская двустволка
- 1.1 История появления ИЖ-27
- 1.2 Особенности конструкции ИЖ-27
- 1.3 Основные технические характеристики ИЖ-27
- 1.4 Модификации ИЖ-27
- 1.5 Ружьё Иж-27М и МР-27М «Юнкер»
- 1.6 Другие популярные модели ИЖ-27
- 1.7 Штучное ружьё ИЖ-27 «Русь»
- 1.8 На какие нюансы нужно обратить внимание при покупке ИЖ-27
- 1.9 ИЖ-27 или ТОЗ-34 – какую двустволку выбрать?
- 2 ИЖ 27
- 2.1 История
- 2.2 Маркировки
- 2.3 Калибры
- 2.4 Стволы
- 2.5 Дульные сужения
- 2.6 Ударно-спусковой механизм ИЖ 27
- 2.7 Эжекторные механизмы
- 2.8 Деревянные детали
- 2.9 Видео о ружье ИЖ 27
ИЖ-27 – самая популярная советская двустволка
Гладкоствольное ружьё ИЖ-27 является обновлённой модификацией модели ИЖ-12, которая в настоящее время практически не встречается. В отличие от ИЖ-12, ИЖ-27 до сих пор очень популярно на просторах СНГ. Новое оружие появилось в 1973 году и через некоторое время стало самой продаваемой двустволкой на территории СССР. Вертикалка ИЖ-27 полюбилась многим охотникам за свои технические характеристики и невероятную надёжность. В те годы охотничье оружие производства Ижевского механического завода собирались очень качественно, в отличие от оружия, которое производится в настоящее время.
В отличие от ИЖ-12, ИЖ-27 до сих пор очень популярно на просторах СНГ. Новое оружие появилось в 1973 году и через некоторое время стало самой продаваемой двустволкой на территории СССР. Вертикалка ИЖ-27 полюбилась многим охотникам за свои технические характеристики и невероятную надёжность. В те годы охотничье оружие производства Ижевского механического завода собирались очень качественно, в отличие от оружия, которое производится в настоящее время.
История появления ИЖ-27
История появления охотничьего ружья ИЖ-27 началась в начале 1970-х годов. Именно в это время группа конструкторов Ижевского механического завода занималась модернизацией двустволки ИЖ-12. Модернизированная модель оружия получила следующие изменения в конструкции:
- Цевье и приклад на ИЖ-27 получили новую форму. Охотники сразу отметили, что ружье стало удобнее;
- Прицельная планка стала вентилируемой;
- Появились перехватыватели курков, которые сделали охоту более безопасной;
- Механический предохранитель, который был установлен на ИЖ-12 заменили автоматическим;
- Появился эжектор;
- Ствольная коробка в месте её соединения с ложей получила новую форму;
- Появился резиновый затыльник на прикладе.

Новая модель успешно прошла все испытания, и в 1973 году оружие было запущено в серию под названием ИЖ-27.
Особенности конструкции ИЖ-27
Вертикальная двустволка выпускается в разных калибрах и имеет следующие особенности конструкции:
- Прицельные приспособление состоят из прицельной планки и мушки;
- Дульные сужения могут быть как постоянными, так и сменными;
- Патронники и ствольные каналы покрыты качественным хромовым покрытием. Снаружи стволы оксидированы;
- Стволы соединяются со ствольной коробкой за счёт подствольных крюков и шарнирной оси;
- Стволы запираются за счёт использования запорной планки, которая заходит в специальный паз заднего подствольного крюка;
- Курки ружья взводятся при открывании стволов;
- Ударно-спусковой механизм имеет возможность произведения спуска курков с боевого взвода, в момент закрывания стволов;
- Гильзы извлекаются из патронников экстрактором;
- Автоматический предохранитель ИЖ-27 одновременно перехватывает курки и запирает шептала.

По сравнению с ИЖ-12, новая модель стала более технологичной и современной.
Основные технические характеристики ИЖ-27
Список основных характеристик выглядит следующим образом:
- Длина ствола двустволки может быть как 750, так и 675 мм;
- Оружие выпускается в 12, 16 и 20 калибрах. Крайне редко встречается модификация под патрон 410 калибра;
- Патронник ИЖ-27 может быть как 70, так и 76 мм. Предпочтительней будет покупка ружья с более длинным патронником, так как она допускает использование патронов с усиленной навеской пороха;
- Масса двустволки составляет от 3,2 до 3,4 кг, и зависит от длины ствола.
Ложа и цевье производится из качественного ореха или бука, традиционно ореховые экземпляры более дорогие. Некоторые модификации имеют в комплекте сменные стволы.
Модификации ИЖ-27
За годы производства было выпущено множество модификаций ИЖ-27. В настоящее время они производятся под индексом МР-27.
- Серийные модели;
- Экспортные модели;
- Штучные модели;
- Сувенирные модели.
Так как перечислить все модификации не представляется возможным, следует остановиться на наиболее интересных и популярных модификациях.
Ружьё Иж-27М и МР-27М «Юнкер»
Модификация ИЖ-27М (сейчас оно выпускается как МР-27М) – это двуствольное ружьё с двумя крючками, и без эжектора. Данная модификация является универсальным охотничьим ружьём, пригодным для охоты на любого зверя. Блок стволов ИЖ-27 имеет калибр 12/76 мм. Благодаря тому, что при изготовлении используется овально-винтовая сверловка, стрельба из этого ружья пулей может вестись на дистанции до 100 метров. Сверловка Ланкастера придаёт пуле дополнительную стабилизацию в полёте за счёт вращения. Для лучшего результата рекомендуется использовать пули «Бреннеке» или «Гуаланди».
При стрельбе картечью или дробью ИЖ-27М обеспечивает неплохую осыпь.
Чтобы добиться лучших показателей при стрельбе дробью, рекомендуется устанавливать сменный ствольный блок. Он имеет открытый прицел, который можно регулировать как по горизонтали, так и по вертикали.
Модель МР-27М «Юнкер» является облегчённой и компактной модификацией ИЖ-27М. Эта двустволка была произведена специально для женщин и подростков. Именно поэтому были изменены размеры её ложи. Так как в России редко кто из охотников станет покупать ружьё для подростка, а женщины традиционно не любят спортивную стрельбу и охоту, то немногие экземпляры МР-27М «Юнкер» применяются опытными охотниками для ходовых охот.
Другие популярные модели ИЖ-27
Кроме ИЖ-27М, в линейке имеются следующие популярные модификации:
- ИЖ-27ЕМ – эта модификация имеет эжектор селективного типа. Он выбрасывает только стреляные гильзы, а целые патроны только выдвигает из стволов. В случае необходимости, данный эжектор можно отключить. Данное ружьё имеет два спусковых крючка и сменные стволы;
- Модель ИЖ-27М-1С – это модификация без эжектора.
 Спусковой крючок у этой модификации только один. Стрельба ведётся поочерёдно, сначала из одного ствола, потом из другого;
Спусковой крючок у этой модификации только один. Стрельба ведётся поочерёдно, сначала из одного ствола, потом из другого; - Модель МР-27Е-1С тоже имеет один спусковой крючок, но в отличие от предыдущей модели, переключение очерёдности стволов происходит за счёт отжима спускового крючка вперёд, а не назад. Это движение долее естественное, поэтому МР-27Е-1С немного более популярно, чем ИЖ-27М-1С.
Российские охотники, привыкшие к наличию двух спусковых крючков у двустволок, предпочитают двухкурковые модификации ИЖ (МР)-27.
Штучное ружьё ИЖ-27 «Русь»
Для ценителей марки «Иж» и модели ИЖ-27 в частности, существует интересная штучная модификация – ИЖ-27 «Русь». Данная модификация имеет две пары дополнительных стволов, серебряную гравировку и отличается улучшенным качеством ручной сборки. Кроме того, подгонка узлов оружия находится на высшем уровне. Цена на это изделие соответствующая: более чем в два раза превосходящая стоимость обычной модели ИЖ-27.
Те же охотники, которые, несмотря ни на что, приобрели данное ружьё, отмечают значительные отличия данной модификации от стандартной модели. Она практически безукоризненно выполнена и превосходит стандартную двустволку по всем параметрам.
Кроме данной модели, непосредственно на заводе можно заказать ИЖ-27 с инкрустациями, использованием экзотической древесины для изготовления ложи и цевья, а также ружьё может быть полностью собрано вручную. Естественно, что за все эти изыски придётся хорошо заплатить.
На какие нюансы нужно обратить внимание при покупке ИЖ-27
При покупке ИЖ-27 (если это современный вариант, то МР-27) можно купить как новое оружие в магазине, так и бывшее в употреблении на вторичном рынке. Если покупать старое ружьё, то основное внимание следует уделить состоянию оружия и работоспособности его механизмов. Часто за небольшую цену удаётся приобрести штучные экземпляры с минимальным настрелом.
При покупке нового ружья желательно иметь возможность выбора из нескольких единиц товара. При выборе нужно обращать внимание на следующие нюансы:
- Кривизна стволов. Иногда производитель выпускает ружья с кривыми стволами. Кроме того, кривые мушки – тоже не редкость при покупке ижевского оружия;
- Пайка стволов. Соединение стволов – очень важная деталь, внимательно осматривайте качество пайки по всей длине соединения. Белый налёт из-под соединительной планки – это повод отказаться от данного экземпляра;
- Дульные срезы стволов должны быть круглой формы, а их толщина – одинаковой. Часто недобросовестные продавцы, пользуясь неопытностью молодых охотников, продают такие ружья, утверждая, что неравномерные дульные срезы – это норма. Как правило, продавцы начинают оперировать такими понятиями, как чок и получок, утверждая, что это доказательство разных дульных сужений;
- Особое внимание нужно обращать на хромировку ствольных каналов ружья.
 В новых моделях МР-27 она иногда бывает неравномерной и имеет пятна;
В новых моделях МР-27 она иногда бывает неравномерной и имеет пятна; - Прицельная планка должна быть ровной. Любые перекосы, выпуклости или углубления на ней – повод отказаться от покупки. Если планка ровная, но на ней присутствуют заусеницы, то это можно быстро устранить своими руками;
- Перед более пристальным осмотром, ружьё нужно просто взять в руки и сильно встряхнуть. Любое дребезжание при этом указывает на неисправность оружия;
- Новое ружьё должно легко открываться и закрываться. Его разборка должна проходить легко, никакие заклинивания недопустимы. Уверения продавца, что всё разработается со временем, следует пропускать мимо ушей;
- Особое внимание нужно обращать на качество подгонки цевья и ложи ИЖ-27 к металлическим частям оружия.
Если сравнивать качество сборки, то можно заметить, что более старые модификации собраны значительно качественней последних, а уж тем более МР-27, поэтому при встрече на вторичном рынке ИЖ-27 в идеальном состоянии, лучше приобрести его, вместо новой двустволки МР-27.

ИЖ-27 или ТОЗ-34 – какую двустволку выбрать?
Если российский охотник хочет приобрести для себя качественную горизонтальную двустволку отечественного производства, то выбор у него всего из двух моделей:
Эти две марки давно уже конкурируют друг с другом, и каждая из них имеет целую армию своих сторонников. Рассмотрим основные достоинства и недостатки этих моделей:
- Качество стволов у ТОЗ-34 традиционно лучше. Если ружьё выбирается для профессионального охотника, который охотится в течение всего года, то для него ресурс стволов будет иметь решающее значение. Для большинства любителей ресурса стволов ИЖ-27 будет более чем достаточно;
- Качество пайки столов у ТОЗ-34 тоже выше, чем у ИЖ-27. У ижевской двустволки чаще отпаивается боковая планка;
- Механизм запирания стволов у двух моделей работает без нареканий и очень надёжен;
- Боевые пружины ИЖ-27 спирального типа, поэтому они крайне редко выходят из строя, а вот ТОЗ-34 этим похвастаться не может.
 Фигурные боевые пружины тульской двустволки очень нежные, поэтому их выход из строя бывает значительно чаще. Так же у ТОЗ-34 бывают проблемы, связанные с выходом из строя рычагов взвода ударников;
Фигурные боевые пружины тульской двустволки очень нежные, поэтому их выход из строя бывает значительно чаще. Так же у ТОЗ-34 бывают проблемы, связанные с выходом из строя рычагов взвода ударников; - Цевье ИЖ-27 часто ломается или трескается. Цевье ТОЗ-34 значительно надёжнее;
- Иж-27 значительно тяжелее изящной двустволки ТОЗ-34. Особенно актуально различие в весе оружия при долгих ходовых охотах;
- ТОЗ-34 более прикладистое.
Что касается надёжности механизмов, то в этом вопросе ИЖ-27 является бесспорным лидером. В итоге получается, что по качеству стволов и общей сборки, ТОЗ-34 значительно лучше. Зато в плане надёжности работы механизмов – ИЖ-27 находится вне конкуренции. Исходя из этого, ТОЗ-34 можно рекомендовать более аккуратным охотникам, которые тщательно следят за своим оружием. ИЖ-27 значительно проще в обслуживании и простит своему владельцу небрежность в эксплуатации, поэтому выбор нужно делать в зависимости от личных предпочтений.
В любом случае, несмотря на то, какую модель выберет для себя охотник, не стоит забывать, что любое оружие нуждается в уходе.
Не стоит пренебрегать чисткой ружья после каждой стрельбы, а также смазкой его механизмов. Если пренебрегать процедурой чистки, скоро внутри стволов появится так называемая «сыпь». Особенно это актуально по отношению к ИЖ-27.
ИЖ-27 или его более современная модификация МР-27 является отличным оружием, надёжность которого проверена десятилетиями. Только вот качество сборки ижевских рядовых ружей не радует покупателя. Часто в магазине приходится выбирать из нескольких десятков МР-27 одно ружьё с нормальным качеством сборки. Если будет возможность приобрести ИЖ-27, которое было выпущено ещё в СССР, и оно будет находиться в хорошем рабочем состоянии, лучше взять именно его. Оно прослужит ещё не один десяток лет.
ИЖ 27
История
Гладкоствольное переломное ружье с вертикально расположенными стволами ИЖ 27 – одно из самых известных отечественных охотничьих ружей, выпускаемое Ижевским механическим заводом с 1972 года. Конструкция ИЖ 27 была создана на базе другого популярного ижевского бокфлинта – ИЖ 12. Начиная с 1976 года группа конструкторов в составе А.А. Климова, Г.Я Протопопова, В.П. Вотякова, Е.М. Губина создала ряд модификаций модели ИЖ 27, в том числе ИЖ 27Е, ИЖ 27-1С, ИЖ 27Е-1С, ИЖ 27ЕМ, ИЖ 27М-1С, ИЖ 27ЕМ-1С.
Начиная с 1976 года группа конструкторов в составе А.А. Климова, Г.Я Протопопова, В.П. Вотякова, Е.М. Губина создала ряд модификаций модели ИЖ 27, в том числе ИЖ 27Е, ИЖ 27-1С, ИЖ 27Е-1С, ИЖ 27ЕМ, ИЖ 27М-1С, ИЖ 27ЕМ-1С.
Маркировки
Маркировка «1С» означает, что модель оснащена одним спусковым крючком, обеспечивающим переключение последовательности выстрелов. Буква «Е» свидетельствует о том, что в ружье установлен эжектор для извлечения гильз. Маркировка «М» — означает длину патронника 76,2 мм, предоставляющую возможность использовать патроны с длиной гильзы 76 мм или патроны «Магнум». С 1972 года и до настоящего времени было изготовлено более миллиона экземпляров ИЖ-27, значительная часть которых экспортировалась за рубеж. Модификации ИЖ 27 Е и ИЖ 27 Е-1С были награждены дипломами и золотыми медалями на международных ярмарках в Лейпциге и Пловдиве.
Калибры
Ружья ИЖ-27 выпускают как серийно, так и в штучном и сувенирном вариантах. Ружья выполнены в 12, 16, 20, 28 и 32 калибрах, а также под патроны «Магнум» — в 12, 20 и . 410 калибрах.
410 калибрах.
Стволы
Стволы у ИЖ-27 съемные, спаяны между собой межствольными планками и скреплены муфтой в казенной части. На верхнем стволе установлена обычная или вентилируемая прицельная планка. Стволы крепятся к колодке при помощи шарнира и подствольных крюков. Стволы запираются рамкой, которая заходит в паз заднего подствольного крюка. Механизм запирания управляется рычагом, находящимся на верхней плоскости коробки. Этот рычаг после открывания стволов удерживается запорной планкой, которая отпускает рычаг при закрывании. Стволы ружей ИЖ 27 производят из прочной ствольной стали, внутренние поверхности стволов, а также патронники хромированы, наружные поверхности оксидированы или хромированы в черный цвет. Длина стволов у ружей 12-го калибра может составлять 675, 725 или 750 мм, длина стволов 16-калиберных ружей равна 725 мм, ружья 20, 28 и 32 калибра выпускают со стволами длиной 675 мм. Длина патронников у ружей всех калибров одинаковая и составляет 70 мм, у ружей под патроны «Магнум» — 76,2 мм. Диаметр канала ствола у ружей равен
Диаметр канала ствола у ружей равен
Дульные сужения
Номинальные значения дульных сужений для 12, 16 и 20 калибров составляют 1 мм для верхнего ствола и 0,5 мм – для нижнего, у ружей 28 и 32 калибров соответственно 0,6 и 0,3 мм, для калибра .410 «Магнум» — 0,5 и 0,25 мм. Отдельные образцы ружей ИЖ-27 12-го калибра могут быть укомплектованы сменными дульными насадками со значениями сужения 1,25; 1,0; 0,75; 0,5; 0,25; 0,0 мм. Перечисленные насадки предназначены для обеспечения постоянства кучности стрельбы. Производитель в прилагаемом к ружью паспорте указывает стандартные значения показателей кучности стрельбы патронами с гильзами длиной до 70 мм, приведенные в нижеследующей таблице.
| Маркировка на стволе | ДР | С | С1 | М | 1М | F | Т |
|---|---|---|---|---|---|---|---|
| Дистанция стрельбы, м | 20 | 20 | 35 | 35 | 35 | 35 | 35 |
| Кучность, %, не менее | 65 | 65 | 40 | 50 | 55 | 60 | 65 |
Ружье ИЖ-27 пригодно для использования патронов с бумажными, пластиковыми и металлическими гильзами, однако при стрельбе патронами с металлическими гильзами кучность боя слегка снижается. Возможно применение как дымного, так и бездымного пороха.
Возможно применение как дымного, так и бездымного пороха.
Ударно-спусковой механизм ИЖ 27
Ударно-спусковой механизм ИЖ-27 расположен в колодке и установлен на отдельном основании. Бойки и курки представляют собой самостоятельные детали. При открывании стволов осуществляется взведение курков и сжатие цилиндрических спиральных боевых пружин. Ударно-спусковой механизм имеет два спусковых крючка и в целях безопасности оснащен автоматическим предохранителем и перехватывателями (интерсепторами) курков. Конструкция ударно-спускового механизма обеспечивает плавный спуск курков. Перехватыватели курков препятствуют выстрелу из-за случайного спуска курков без нажатия на спусковые крючки. Для длительного хранения ружья необходимо разгрузить боевые пружины путем двукратного нажатия на спусковые крючки. Уружей с маркировкой «1С» единственный спусковой крючок обеспечивает следующуюпоследовательность выстрелов: при первом нажиме происходит выстрел из нижнегоствола, при втором нажиме – из верхнего. Последовательность на очередные двавыстрела можно изменить за счет отжима спускового крючка вперед до щелчка. Приочередном заряжании ружья исходная последовательность выстрелов восстановится автоматически.
Последовательность на очередные двавыстрела можно изменить за счет отжима спускового крючка вперед до щелчка. Приочередном заряжании ружья исходная последовательность выстрелов восстановится автоматически.
Эжекторные механизмы
У ружей в стандартном исполнении гильзы из патронников извлекаются экстрактором. Автоматический эжекторный механизм ружей ИЖ-27, маркированных буквой «Е», извлекает только отстрелянные гильзы, но при необходимости его можно отключить и устройство будет выполнять функции экстрактора. Эжекторные механизмы этих ружей требуют аккуратного обращения. Ошибки при разборке и сборке ружей, особенно с приложением больших усилий, могут привести к деформациям деталей эжекторов, что неминуемо приведет к необходимости квалифицированного ремонта.
Деревянные детали
Деревянные детали ружей ИЖ-27 изготавливают из ореха или бука, цевье выполнено съемным, закрепляющимся с помощью защелки. На ружья устанавливают пластиковый затыльник или резиновый амортизатор.
Видео о ружье ИЖ 27
Post Views: 75
Подбор гильз и патронов для ИЖ-27
В 1986 году в магазине я приобрел штучное ружье ИЖ-27, с паспортом и печатью мастера-штучника Самарина Николая Васильевича и со знаком качества.
Фото: Евгений Копейко.
Данные ружья по паспорту: длина стволов — 723 мм; диаметр каналов стволов 18,2 мм; патронник — 70 мм, верхний ствол — чок, нижний ствол 0,5 — получок.
Охочусь я с 1976 года. Каково было мое удивление, что после стрельбы из ружья магазинными бумажными патронами была сильная отдача в плечо и до крови были разбиты пальцы на руке. Выход из положения — была уменьшена навеска пороха и дроби до разумной стрельбы.
Так я охотился с этим ружьем несколько лет. Но однажды в газете «РОГ» прочитал статью, как один пожилой охотник при встрече с охотниками у костра на привале увидел у одного из них перебинтованные пальцы рук и попросил показать ему ружье, с которым тот охотился. Ружье было ИЖ-27, «Штучное».
Ружье было ИЖ-27, «Штучное».
Всю причину большой отдачи ружья пожилой охотник-токарь мне продемонстрировал: достав из кармана деревянный макет патрона длиной 76 мм и вставив его в патронник, он спокойно закрыл стволы. Точно такое же ружье было у пожилого охотника-токаря, «ИЖ-27», штучное.
Я дал молодому свои 76 мм патроны для пробы пострелять; отдача была мягкая и намного меньше против 70 мм патронов!
Каково же было мое удивление, когда я вставил в патронники новые пустые 76 мм гильзы, легко закрыл стволы. После опробования стрельбой такими гильзами с навеской пороха «Сокол» 2,3 г, дроби 35 г, я увидел и ощутил прекрасный результат во всем.
После замера диаметра канала стволов за переходным конусом он оказался не 18,2 мм по паспорту, а 18,4 мм, геометрия нижнего ствола — 0,8 мм, геометрия верхнего ствола — 1,1–1,2 мм.
Я с уважением отношусь к консультанту Евгению Геннадьевичу Копейко. Являюсь его постоянным читателем и учеником. Пожалуйста, если можно, как он, большой знаток оружия, прокомментирует это, и как правильно понять и эксплуатировать это оружие, с 76 мм гильзами или с 73 мм гильзами, так как 76 мм новая пустая чуть-чуть не заходит до конца, а при закрытии стволов затруднений нет никаких.
Пожалуйста, если можно, как он, большой знаток оружия, прокомментирует это, и как правильно понять и эксплуатировать это оружие, с 76 мм гильзами или с 73 мм гильзами, так как 76 мм новая пустая чуть-чуть не заходит до конца, а при закрытии стволов затруднений нет никаких.
Хочется также от себя и моих товарищей выразить большую благодарность Анатолию Азарову — охотнику от Бога за его статьи, опубликованные неоднократно в «РОГ».
П.Н. КОВАЛЬ
Гильзы и патроны для вертикалки ИЖ-27 12-го калибра с патронниками длиной 70 мм
| |
Фото Антона Журавкова. |
Вопрос применения гильз длиной 70 мм или любой другой длины в гладкоствольных ружьях с патронниками длиной 70 мм решен давным-давно.
Суть заключается в соответствии патронов и гильз патроннику и стволу охотничьего ружья. Но если такой вопрос возник, то придется ответить подробнее, так как несоответствие может быть опасно.
Принимать во внимание необходимо многое, но пока ограничимся геометрическими параметрами ружейного ствола, гильз и патронов, а также допускаемым для ружья давлением.
Вертикалок ИЖ-27 с различными параметрами много, есть экземпляры высокой категории, как на фотографии Копейко Е.Г.. Но речь пойдет о ружье ИЖ-27 12-го калибра с патронниками длиной только 70 мм, как в письме автора П.Н. Коваля.
Каналы стволов дробовых ружей имеют вполне определенные размеры, регламентированные правилами международной комиссии C.I.P. (ПМК в России). Так для ружья 12-го калибра принят диаметр канала ствола в пределах 18,20–18,90 мм. Измеряется диаметр канала ствола в определенной точке — на расстоянии не более 220 мм от казенного среза, обычно 150–220 мм. Эти пределы учитывают все производители оружия.
Так и ваше ружье ИЖ-27 «Штучное» 1986 года оснащено блоком стволов с диаметрами каналов, равными 18,2 мм, как следует из вашего указания на «Паспорт ружья».
Разница паспортного значения и измеренного диаметра 18,4 мм довольно велика для стволов, изготовленных методом холодной ковки, но может быть объяснена производственными допусками, если измерения проведены и на указанном выше расстоянии от казенного среза, и до стрельбы патронами с гильзами длиной 76 мм.
Требования к размерам патронников гладкоствольных ружей намного строже, так диаметр патронника 12-го калибра у казенного среза ствола должен быть в пределах 20,65–20,75 мм.
Патронники конические, сужаются по направлению к переходному конусу в канал ствола, где диаметр патронника 12-го калибра составляет 20,30–20,40 мм. Обращаю на это внимание, так как это одна из основ конструирования гладкого ствола охотничьего ружья.
Причина ясна — патронник должен быть универсальным, чтобы применять патроны любых производителей.
Соответственно патронникам регламентируются и размеры гильз. Так для гильз 12-го калибра строго устанавливается наружный диаметр: у ранта 20,32–20,60 мм, у дульца 19,73–20,20 мм.
Наружные размеры любых гильз 12-го калибра должны укладываться в эти пределы.
А внутренние диаметры гильз так строго не регламентируются, они зависят от материала обечайки гильзы, который должен выдерживать высокое давление и температуру при выстреле.
Изготавливаются они из бумаги — самого распространенного материала, весьма технологичного пластика, набирающего все большую популярность и быстро вытесняющего остальные виды, а также из металла, в частности латуни, стали, алюминия.
Наибольшая толщина стенок у бумажных гильз, самые тонкостенные это металлические, а пластмассовые по толщине стенок занимают промежуточное положение между ними. Как правило, внутренний диаметр бумажных гильз 12-го калибра лежит в пределах 18,3–18,6 мм, а пластмассовых в диапазоне 18,5–18,8 мм. Металлические гильзы я не рассматриваю, они — в прошлом.
Идеальное сочетание гильзы и ствола охотничьего ружья заключается в совпадении внутреннего диаметра гильзы у дульца (далее по тексту — внутреннего диаметра гильзы) и диаметра канала ствола. Но так практически не бывает, точнее, бывает чрезвычайно редко.
Разница между диаметром канала ствола и внутренним диаметром гильзы при переходе снаряда дроби из гильзы в канал ствола и возникший при этом скачок давления и есть одна из основных причин отдачи.
Чем больше эта разница, тем сильнее отдача. Явление естественное, избежать его невозможно, а ослабить его действие можно, подбирая толстостенные гильзы.
Очень многие двустволки 12-го калибра XX века, отечественные, германские, французские, имеют стволы с каналами диаметром 18,2–18,3 мм, и охотники давно научились подбирать к ним нужные гильзы.
Когда-то в охотничьем обиходе были такие понятия, как «ствол, сверленный под металлическую гильзу», и «ствол, сверленный под бумажную гильзу», конечно, в пределах одного и того же калибра.
Несоответствие гильзы и ствола отрицательно влияло на показатели боя дробью, хотя, справедливости ради, отмечу, что стрельба из ружья с широкими каналами стволов практически не сказывалась на отдаче.
При стрельбе пальцы действительно могут быть повреждены спусковыми крючками, обычно передним, как в результате отдачи, так и при неправильном, точнее неловком положении ладони стрелка на шейке ложи. Руки у охотников разные, а шейка ложи типовых серийных ружей стандартная, рассчитанная на стрелка осредненного телосложения.
Индивидуальная ложа стоит немалых денег, ложевщик должен снять с охотника несколько размеров и воплотить их в древесине заготовки, но она оправдана при частой стрельбе.
Для защиты пальцев на ружья высокого класса устанавливают спусковые механизмы с подпружиненным передним спусковым крючком. Удобен и механизм с одним спусковым крючком.
А теперь о самом главном, о длине гильзы и длине патронника. Понятно, что гильза в патроннике большей длины, чем она сама, приведет только к ухудшению показателей боя дробью за счет прорыва пороховых газов в зоне между кромкой дульца гильзы и началом переходного конуса.
Для ружья это не опасно, хотя упустить дичь по такой нелепой, легко устраняемой причине весьма и весьма досадно.
Гораздо опаснее гильза длиной 76 мм в патроннике меньшей длины 70 мм. Ее дульце, обжатое в переходном конусе, уменьшает свободную площадь поперечного сечения и вызывает местный скачок давления, способный вызвать раздутие ствола.
Ремонтировать раздутый за патронником ствол бесполезно, последующие выстрелы приведут к разрыву. Именно поэтому так тщательно маститые охотники рассматривают стволы старых ружей, чтобы обнаружить раздутие, шустовку каналов и наружную опиловку.
При веском подозрении такие ружья, даже ценные, бракуются, безопасность охотника важнее «удачной» покупки. За все время существования казнозарядных охотничьих ружей и унитарных патронов в результате такой стрельбы погибло колоссальное количество ценных охотничьих ружей, и только потому, что охотники не удосужились подобрать патроны по патронникам.
Патрон, снаряженный в гильзу длиной 76 мм, поместится в патронник длиной 70 мм, так как гильза укорочена на длину завальцованной части дульца. Но при выстреле дульце раскроется как раз в переходном конусе и вызовет скачок давления.
Но при выстреле дульце раскроется как раз в переходном конусе и вызовет скачок давления.
Если повезет, ствол от одного выстрела не раздуется. Но в этом месте на него подействовало высокое давление, и при последующих выстрелах раздутие обнаружится.
В патронник длиной 70 мм можно вставить пластиковую или бумажную гильзу длиной 76 мм и закрыть блок стволов, действительно дульце обомнется в переходном конусе под небольшим усилием. Создается ложное впечатление, что такая гильза пригодна, но в этом как раз и заключается опасность раздутия ствола.
Еще один важнейший аспект выбора патронов заключается в соответствии давления, развиваемого патроном при выстреле, допускаемом для ружья давлению, оно указывается на блоке стволов и документации.
Из вертикалок ИЖ-27 12-го калибра с патронниками длиной 70 мм можно стрелять только патронами с гильзами длиной 70 мм, развивающими давление до 72 МПа. А более мощные патроны с гильзами длиной 76 мм, развивающие давление до 90 МПа, вообще не для них.
Вертикалка ИЖ-27 «Штучное» 12-го калибра 1986 года от мастера Самарина Николая Васильевича — образец весьма достойный, надолго переживет многие нынешние иноземные ружья, а потому заслуживает уважения и хорошо подобранных патронов с гильзами длиной 70 мм.
Задать свой вопрос Евгению Копейко можно по электронному адресу: [email protected]
Евгений Копейко 28 марта 2017 в 00:02
Ружье ИЖ-27 характеристики и виды
История создания
Разработка рассматриваемой модели была начата в конце 60-х. Группе проектировщиков во главе с А. А. Климовым была поставлена задача устранить недостатки выпускающейся в то время модели ИЖ12. В 1971 году инженеры представляют первый опытный образец. 1973 – год выпуска ИЖ-27 в массовое производство.
Неизменным осталось расположение стволов один над другим. Во всем же остальном конструкция оружия претерпела серьезные изменения. Прежде всего, это более удобная и эргономичная, в сравнении с ИЖ-12, форма приклада и цевья. Конструкторы установили рычаг, перехватывающий курок при случайном срыве шептала с курка, а также авто предохранитель, сделали отверстия в прицельной планке (для обеспечения вентиляции). Было изменено место соединения приклада с затворной коробкой. Установленный на него резиновый затыльник эффективно снижает отдачу при выстреле и ее воздействие на стрелка. Но самое главное, ИЖ-27 – первая модель ИМЗ с эжектором для гильз.
Конструкторы установили рычаг, перехватывающий курок при случайном срыве шептала с курка, а также авто предохранитель, сделали отверстия в прицельной планке (для обеспечения вентиляции). Было изменено место соединения приклада с затворной коробкой. Установленный на него резиновый затыльник эффективно снижает отдачу при выстреле и ее воздействие на стрелка. Но самое главное, ИЖ-27 – первая модель ИМЗ с эжектором для гильз.
Ружье гладкоствольное ИЖ-27ЕМ
ИЖ-58 — Википедия
Материал из Википедии — свободной энциклопедии
| ИЖ-43 | |
| Тип | двуствольное ружьё |
| Страна | СССР |
| Конструктор | |
| Производитель | Ижевский механический завод |
| Годы производства | с 1958 до 1980х |
| Масса, кг | 2,7-2,9 (20-й калибр)3,2 (12-й и 16-й калибр) |
| Длина ствола, мм | 675, 720, 730 |
| Калибр | 12/70, 16/70, 20/70, 28/70 |
ИЖ-58
— советское двуствольное охотничье ружьё.
Ружьё ИЖ-58 было разработано в 1958 году в качестве лёгкого ружья 20-го и 28-го калибра для промысловой охоты, ходовой охоты в тайге и охоты в горах во всех климатических зонах СССР, первые ружья были изготовлены в конце 1958 года
.
С 1960 года начался выпуск ружей ИЖ-58 под патрон 16-го калибра, на базе которого в 1970 году было создано ружьё ИЖ-58М под патрон 12-го калибра (серийный выпуск которого начался в 1971 году).
Ружья 28-го калибра серийно не выпускались, ружья 20-го калибра выпускались с конца 1958 года до середины 1960-х годов, а основная часть ИЖ-58 была выпущена под патроны 12-го и 16-го калибра. Всего было выпущено свыше 1,36 млн. ружей ИЖ-58 всех четырёх модификаций. Кроме того, было изготовлено опытное ружьё ИЖ-58 20-го калибра с алюминиевой коробкой, армированной стальным штифтом и стволами без соединительных планок, соединёнными тремя муфтами.
В 1980е годы началась разработка нового ружья на замену ИЖ-58 и его модификаций, которым стало ИЖ-43.
ИЖ-58 представляет собой двуствольное ружьё с горизонтальным расположением стволов. Трубки стволов и казённая муфта изготавливались из стали 50А. Стволы отъёмные, имеют параболические дульные сужения.
- дульное сужение правого ствола – 0,5 мм (получок)
- дульное сужение левого ствола – 1 мм (полный чок)
Патронники и каналы стволов хромированы.
Запирание ружья тройное: на два нижних крюка запорной планкой и на верхний крюк – рычагом запирания.
Спусковой механизм с двумя спусковыми крючками смонтирован на отдельном основании, усилие спуска – в пределах от 1,5 до 2,5 кг. Боевые пружины – винтовые цилиндрические, они взводятся шарниром цевья и толкателями при открывании ружья. Бойки диаметром 2,5 мм впрессованы в курки и имеют механизм отбоя.
Предохранитель неавтоматический, запирает шептала (в случае срыва курков с боевого взвода они автоматически встают на предохранительный взвод, не ударяя по капсюлям). При не полностью закрытых стволах выстрела не происходит.
Колодка ИЖ-58 изготавливалась из нормализованных поковок прочной конструкционной стали 50 (не нуждавшейся в последующей цементации) с последующей термообработкой и чистовой доводкой базовых размеров и поверхностей на специализированных станках.
Приклад и цевье изготавливают из берёзы, бука или (на штучных ружьях) – из ореха. Цевье отъёмное, закреплено на межствольной планке защёлкой. Ложа прямая или пистолетная
.
Ружьё отличается простой и технологичной конструкцией, его полная разборка проводится без применения специальных инструментов (нужны только выколотка и отвёртка).
- ИЖ-58
- ИЖ-58М
– ружьё 12-го калибра образца 1970 года - ИЖ-58МА
– модель с новым автоматическим предохранителем, созданная в середине 1970-х годов. Гарантированная отстрелом живучесть ИЖ-58МА составляет 15 тысяч выстрелов - ИЖ-58МАЕ
– модель с новым автоматическим предохранителем и эжектором, выпуск которой начался в 1977 году
- ↑ 1234567891011121314151617181920212223
А. В. Кузьминский. Оружие для охотника: практическое пособие / под общ. ред. А. Е. Тараса М., ООО «Издательство АСТ», 2002. стр.171-173
В. Кузьминский. Оружие для охотника: практическое пособие / под общ. ред. А. Е. Тараса М., ООО «Издательство АСТ», 2002. стр.171-173 - ↑ 123456789
В. Вальнев, Ю. Маслов. ИЖ-58 – 50 лет в строю (часть 2) // журнал “Мастер – ружьё”, № 11, ноябрь 2008 - ↑ В. Вальнев, Ю. Маслов. ИЖ-58 – 50 лет в строю (часть 1) // журнал “Мастер – ружьё”, № 10, октябрь 2008
- ↑ В. Вальнев, Ю. Маслов. Наши ружья // журнал “Мастер – ружьё”, № 2 (71), февраль 2003. стр.12-15
- Universal/Baikal Model IJ-58M shotgun // журнал “American Rifleman”, January 1977
- М. М. Блюм, И. Б. Шишкин. Охотничье ружьё. М., «Лесная промышленность», 1983. стр.85
- Ижевское оружие. Том 1. Ижевские ружья / Н. Л. Изметинский, Л. Е. Михайлов. – Ижевск, издательство Удмуртского университета, 1995. – 247 стр. : ил.
Чем отличается ИЖ-27 от МР-27?
Изначально оружие, выпускаемое Ижмехом, маркировалось двумя буквами ИЖ и цифрами, в соответствии с годом выпуска. В 60-е годы прошлого века цифры в названии оружия стали обозначать не год выпуска, а номер проекта.
В 60-е годы прошлого века цифры в названии оружия стали обозначать не год выпуска, а номер проекта.
В 2008 году, с ростом популярности модели и решением выхода на международный рынок, возникла острая необходимость в переименовании старого ИЖ-27, что связано с неудобством транскрибирования обозначения «ИЖ» на английский язык. Перед руководством завода стояла задача подобрать обозначение, отражающее смысл, и при этом имеющее одинаковое звучание и графику как на латинице, так и на кириллице. И после нескольких мозговых штурмов топ-менеджеров компании такая бренд-аббревиатура была придумана – Mehanical Plant (механический завод). Начиная с того времени и по сегодняшний день, все новые разработки маркируются буквами МР. Соответственно, ИЖ-27 старого образца – это оружие, выпущенное до 2008 года. Современные же модели вместо ИЖ-27 имеют обозначение МР-27.
Обязательные приемочные клейма.
Клеймо — это отдельный знак, отличный от маркировки. Если маркировка несет в себе информацию непосредственно о ружье, то клеймо подтверждает, что это ружье прошло испытание и соответствует предъявляемым требованиям, в первую очередь требованиям безопасности.
2.1.Клеймо проверки прочности оружия усиленными выстрелами.
На сегодняшний день такое клеймо существует в двух видах. Буква П, вписанная в круг, предназначена для гладкоствольного охотничьего оружия (кроме ружей под патрон «Магнум»). Та же буква П, но в двух концентрических кругах — для охотничьих гладкостволок под патрон «Магнум» и спортивного гладкоствольного оружия, так как для второго типа оружия применяется более усиленное испытание. Обычай использования этого клейма имеет, по-видимому, очень старые корни. Буква П или П в круге, означающая проверочное клеймо «Пробован» ставилась еще на клинках Златоустовской оружейной фабрики в 1837-1905 годах, а в Туле, по всей видимости, еще задолго до того. (см. Кулинский А.Н. «Русское холодное оружие военных, морских и гражданских чинов 1800-1917 годов». СП 1994, «Магик-ПРЕСС»,»ОЛИМП»).
На ижевских ружьях это клеймо стало применяться с начала 1984 года с введением отраслевого стандарта на маркировку, на тульских встречалось и ранее, по крайней мере, на ТОЗ-34 оно ставилось ещё в 1970-х годах и означало тогда усиленное испытание стволов. Вместе с ним ставились клейма в виде букв К и У в кругах — подтверждающие испытания соответственно на кучность и меткость, и на усиленное испытание ружья, а также клеймо ОТК завода в виде соединенных букв Т и К в круге. До 1984 года в Ижевске ставились другие аналогичные клейма, например, на ИЖ-49 клеймо испытания ружья усиленными выстрелами выглядело как буква У в трапеции («усиленное испытание»). Позже буква У была заменена буквой Н («нитроиспытание»). Сначала каждый отстрелянный ствол двустволок клеймился словом «НИТРО», затем появилось отдельное клеймо — буква Н в ромбе. Комбинированные «Белки» и одноствольные ружья клеймились буквой Н в круге — видимо испытания гладкого ствола и ружья были совмещены. С 1980 года, по аналогии с клеймами на двуствольных ружьях, на одноствольных тоже вводится два клейма для подтверждения усиленного отстрела ствола и отдельно ружья — буквы Н в ромбе и трапеции.
Вместе с ним ставились клейма в виде букв К и У в кругах — подтверждающие испытания соответственно на кучность и меткость, и на усиленное испытание ружья, а также клеймо ОТК завода в виде соединенных букв Т и К в круге. До 1984 года в Ижевске ставились другие аналогичные клейма, например, на ИЖ-49 клеймо испытания ружья усиленными выстрелами выглядело как буква У в трапеции («усиленное испытание»). Позже буква У была заменена буквой Н («нитроиспытание»). Сначала каждый отстрелянный ствол двустволок клеймился словом «НИТРО», затем появилось отдельное клеймо — буква Н в ромбе. Комбинированные «Белки» и одноствольные ружья клеймились буквой Н в круге — видимо испытания гладкого ствола и ружья были совмещены. С 1980 года, по аналогии с клеймами на двуствольных ружьях, на одноствольных тоже вводится два клейма для подтверждения усиленного отстрела ствола и отдельно ружья — буквы Н в ромбе и трапеции.
2.2. Клеймо Государственной испытательной станции, как организации надзирающей за испытаниями оружия в стране, введено в 1966 году. Оно имело буквы ГИС, очерченные в ромбе. Фактически на станции проводились только приемочные и ежегодные периодические испытания одного ружья каждой модели, включающие в себя отстрел на гарантированную живучесть. После введения обязательной сертификации клеймо ГИС заменено на Знак обязательной сертификации и Знак испытательной станции предприятия (для механического завода г. Ижевска — стрела в круге). На ружье ИЖ-27М это было сделано примерно в октябре 1993 года.
Оно имело буквы ГИС, очерченные в ромбе. Фактически на станции проводились только приемочные и ежегодные периодические испытания одного ружья каждой модели, включающие в себя отстрел на гарантированную живучесть. После введения обязательной сертификации клеймо ГИС заменено на Знак обязательной сертификации и Знак испытательной станции предприятия (для механического завода г. Ижевска — стрела в круге). На ружье ИЖ-27М это было сделано примерно в октябре 1993 года.
После создания Удмуртского центра сертификации, с декабря 1996 года вместо этих двух знаков наносится Знак Удмуртского Государственного Центра в виде стрелы с крылышками в овале, который является признанным клеймом Брюссельской Конвенции. Он наносится только на испытанное усиленным отстрелом оружие.
Особенности МР-27 (ИЖ-27)
- 1. Стволы ИЖ-27 изготовлены из высококачественной оружейной стали и имеют эффективную защиту от коррозии.
- 2. Возможность применения охотничьих патронов с утопленной пулей за счет увеличенной длины бойков.

- 3. Для объединения корпуса оружия и стволов используются крюки и шарнирный механизм.
- 4. Курки плавно и мягко спускаются с боевого взвода сразу же при закрытии стволов, что обеспечивает безопасность эксплуатации. Для достижения этой цели инженеры ИМЗ разработали особую конструкцию ударно-спускового механизма.
- 5. В ИЖ-27 установлен авто предохранитель с перехватывателями курков. Блокируя шептала, он исключает возможность случайного выстрела, который может произойти, например, при сильной тряске в автомобиле или при падении оружия.
- 6. МР-27 может быть оснащено стандартным механизмом для удаления гильз или же выбрасывающим исключительно отстреленные гильзы селективным эжектором. Благодаря этому в случае вылета пули только из одного (верхнего или нижнего) ствола второй патрон остается на своем месте, что позитивно сказывается на времени перезарядки. Эжектор можно отключить.
- 7. Любая из модификаций ИЖ-27 поддерживает применение патронов с длиной гильзы 76 мм.
 Конструкция ружья достаточно прочная, чтобы выдерживать такие мощные боеприпасы.
Конструкция ружья достаточно прочная, чтобы выдерживать такие мощные боеприпасы.
Ружье гладкоствольное ИЖ-27Е
[править] Последовательность выстрелов
Основная последовательность — нижний ствол, затем верхний ствол. Для изменения этой последовательности на противоположную необходимо отжать спусковой крючок вперед до щелчка. Однако такое переключение действует только на одну пару выстрелов, и при первом же открывании ружья основная очередность восстанавливается.
Если после изменения обычной последовательности вдруг возникает необходимость вновь изменить обратную очередность на прямую, необходимо повернуть рычаг запирания стволов вправо и, не открывая стволов, вернуть его в прежнее положение.
Если при нажатии на спусковой крючок произошла осечка, переключение произойдет независимо от того, произошел ли первый выстрел после спуска или нет.
Разновидности ИЖ-27
Линейка оружия с 2 спусковыми крючками представлена моделями МР-27-М и МР-27Е-М. В них каждый спусковой крючок отвечает за выстрел из конкретного ствола. ИЖ-27ЕМ – оружие с эжектором, автоматически сбрасывающим отработанные гильзы. Модификация же ИЖ-27М оснащена экстрактором.
В них каждый спусковой крючок отвечает за выстрел из конкретного ствола. ИЖ-27ЕМ – оружие с эжектором, автоматически сбрасывающим отработанные гильзы. Модификация же ИЖ-27М оснащена экстрактором.
Однокрюковые МР-27 представлены моделями И-Ж-27-М1-С и И-Ж-27-ЕМ-1-С с экстрактором гильз и эжектором соответственно. Огонь ведется последовательно, сначала из верхнего, затем из нижнего ствола, или наоборот. МР-27 ЕМ-1С – модификация для спортивной стрельбы. Ведь эжектор и один спусковой крючок позволяют по максимуму отбросить все лишнее: спортсмену не нужно думать, на какой спусковой крючок нажать, гильза автоматически выбрасывается. Все это позволяет лучше сконцентрироваться, сэкономить драгоценные секунды за счет высокой скорости перезарядки.
Ружье гладкоствольное ИЖ-27ЕМ-1С
Исполнения ИЖ 27 (МР 27)
- 1. Под брендом «Байкал» производитель выпускает, начиная с 2009 года, классическую версию оружия.
- 2. ИЖ-27 М Junior (МР-27М Junior) – облегченная укороченная версия с меньшим калибром.
 Она разработана специально для низкорослых охотников.
Она разработана специально для низкорослых охотников. - 3. ИЖ-27 СТ (снята с производства) – оружие для спортивной стрельбы по летающим тарелкам. Модель имеет самую большую длину ствола из всей серии ИЖ-27 – 760 мм.
- 4. ИЖ-27 СК – также спортивная разновидность, предназначенная уже для скита. Длина стволов в этой модели – 660 мм.
- 5. МР-27 Sporting – одна из последних разработок оружейного завода, выгодно отличающаяся своей универсальностью.
Помимо этого, существуют лимитированные выпуски. Например, ИЖ-27 «Русь». Коллекционные ИЖ-27 легко узнать по серебряной отделке с глубокой гравировкой и интересным рельефным рисунком – изображением различных сцен охоты. Одним из немногих обладателей эксклюзивной модификации «Русь» является наш президент.
Длины стволов ИЖ 27
Стандартная, классическая длина ружья ИЖ 27 –725 мм. Она считается универсальной, подходящей для различных стилей стрельбы. В удлиненных стволах эта характеристика составляет 750 мм (75 см). По сути, 25 мм – это не так уж и много, менее 1 дюйма. Но даже такое незначительное удлинение существенно улучшает поражающие способности боеприпасов: за счет того, что пуля дольше движется по стволу, улучшается разгон, соответственно, и резкость выстрела. Эта характеристика указывает на пробивную силу дроби в момент поражения цели. Для ее измерения нужно взять дробь 3 мм и произвести выстрел в обычную сосновую доску. После этого штангенциркулем либо градуированным щупом измеряется глубина попадания. Если нет под рукой указанных измерительных инструментов, можно воспользоваться спичкой, зубочисткой, иголкой и другими подручными средствами: отметить, на какую глубину утапливается спичка, после чего замерить расстояние от ее края до метки. Чем глубже проникла пуля в доску, тем более резким считается ружье.
По сути, 25 мм – это не так уж и много, менее 1 дюйма. Но даже такое незначительное удлинение существенно улучшает поражающие способности боеприпасов: за счет того, что пуля дольше движется по стволу, улучшается разгон, соответственно, и резкость выстрела. Эта характеристика указывает на пробивную силу дроби в момент поражения цели. Для ее измерения нужно взять дробь 3 мм и произвести выстрел в обычную сосновую доску. После этого штангенциркулем либо градуированным щупом измеряется глубина попадания. Если нет под рукой указанных измерительных инструментов, можно воспользоваться спичкой, зубочисткой, иголкой и другими подручными средствами: отметить, на какую глубину утапливается спичка, после чего замерить расстояние от ее края до метки. Чем глубже проникла пуля в доску, тем более резким считается ружье.
Ружье ИЖ-27 ЕМ
В целом, МР-27 – оружие, которое само по себе имеет хорошую резкость. Модификации с ручной перезарядкой не расходуют пороховые газы на смену заряда – вся мощность направлена в выстрел. Соответственно, и резкость выстрела здесь будет больше. И в этом существенный плюс оружия этой группы.
Соответственно, и резкость выстрела здесь будет больше. И в этом существенный плюс оружия этой группы.
Помимо стволов средней длины и удлиненных моделей, существуют и ИЖ-27 с короткими стволами, так называемые «короткостволки»: 610, 660, 675 мм, а также укороченные ИЖ – 710 мм. Такое оружие используется для ближней охоты, для ходовых работ. Например, для загонной охоты, где расстояние до мишени составляет 15–30 метров, для мелкой дичи, птиц и водоплавающих.
Сверловка ИЖ 27 (МР 27)
Оружие ИЖ-27 предлагается в модификациях с постсверловкой или же с 3 дульными насадками (цилиндр, чок, получок) в комплекте (имеют в маркировке приставку д/н). Модификация д/н подойдет опытным охотникам. Использование разных комбинаций дн дает охотнику возможность менять площадь рассеивания дроби.
В модификациях без сменных дульных насадок применяется классическая сверловка: верхний ствол – чок, нижний – получок. Все выпускаемые ИМЗ сужения унифицированы.
Из каких материалов может быть изготовлен приклад в ИЖ-27
Изначально для изготовления ИЖ-27 применялись исключительно дерево и металл. Пластиковые приклады стали использоваться совсем недавно. Впервые пластиковые приклады появились в военном оружии, для которого на первом месте всегда находится не эстетическая составляющая, а практичность и устойчивость к износам. Пластик упрощает обслуживание оружия. Он не впитывает влагу, не подается гниению, воздействию плесени и грибковых спор, намного проще в уходе. Из существенных недостатков можно отметить только сложности при эксплуатации в условиях низких температур: так как пластик холодный материал, в зимнюю стужу он будет сильно леденить лицо.
Пластиковые приклады стали использоваться совсем недавно. Впервые пластиковые приклады появились в военном оружии, для которого на первом месте всегда находится не эстетическая составляющая, а практичность и устойчивость к износам. Пластик упрощает обслуживание оружия. Он не впитывает влагу, не подается гниению, воздействию плесени и грибковых спор, намного проще в уходе. Из существенных недостатков можно отметить только сложности при эксплуатации в условиях низких температур: так как пластик холодный материал, в зимнюю стужу он будет сильно леденить лицо.
Ижевский завод использует различные сорта древесины. Естественно, пиломатериалы высшего сорта, элитные породы древесины применяются в лимитированных премиум-исполнениях. Дерево – теплый, природный материал. Он не холодит щеку, придает ружью особый шарм, но требует специализированного ухода.
Ружье ИЖ-27 ЕМ-1С
Если вам не по душе холодный и бездушный пластик, но при этом смущает «беззащитность» древесины, стоит присмотреться к исполнению Park. Oil P. Здесь буковая древесина обрабатывается специальным защитным составом, за счет чего существенно улучшаются показатели влагостойкости, прочности, устойчивости к сколам и царапинам, воздействию у/ф лучей. Дополнительным бонусом станут улучшенные визуальные свойства: дерево облагораживается, получает более темный цвет.
Oil P. Здесь буковая древесина обрабатывается специальным защитным составом, за счет чего существенно улучшаются показатели влагостойкости, прочности, устойчивости к сколам и царапинам, воздействию у/ф лучей. Дополнительным бонусом станут улучшенные визуальные свойства: дерево облагораживается, получает более темный цвет.
Впрочем, если вы не можете договориться с собой и решить, что же лучше: дерево или пластик, можно остановиться сразу на двух вариантах! Ведь в МР-27 приклады можно менять!
Прочие надписи и знаки.
Поскольку информация о технологических маркировках крайне скудна и не столь интересна, остается рассказать о знаках, обязательных только в некоторых случаях и о необязательных знаках и надписях.
Обычно в эту группу входят названия торгующих фирм, Знак Качества, надписи «Сделано в России», монограммы владельцев и дарственные надписи. Часть дополнительной маркировки является обязательной только для ружей внутреннего рынка. Другая часть дополнительной маркировки является обязательной только для экспорта, но, поскольку отраслевой стандарт разрешает её наличие для ружей внутреннего рынка, то эта маркировка ставится на все ружья – потому что так проще изготавливать инструмент и не надо вести лишний учет. Конечно, Ижмехзавод после выхода на внешний рынок также старался ввести международную маркировку на всё свое оружие. Тогда же практически перестали выпускать ружья 16-го калибра. Туляки в этом отношении всегда ориентировались на глубинную Россию и Сибирь, выпуская, например, в относительно больших количествах ТОЗ-34 28 калибра.
Конечно, Ижмехзавод после выхода на внешний рынок также старался ввести международную маркировку на всё свое оружие. Тогда же практически перестали выпускать ружья 16-го калибра. Туляки в этом отношении всегда ориентировались на глубинную Россию и Сибирь, выпуская, например, в относительно больших количествах ТОЗ-34 28 калибра.
4.1. Знак Качества по ГОСТ 1.9-67 присваивался разным моделям ружей в разные годы, например на ИЖ-26Е он стал наноситься с 1970 года только на ружья, предназначенные для внутреннего рынка страны. На ружьях экспортного исполнения, проданных в Союзе, этот знак отсутствует. Отменён в конце 1990 года.
4.2. Надпись «Сделано в СССР» наносилась только на коробки ружей экспортного исполнения, обычно на русском языке или, по требованию заказчика, на английском. В марте 1992 года надпись логично трансформировалась на «Сделано в России». На ружьях для внутреннего рынка может отсутствовать.
4.3. Вероятно стоит упомянуть, что фирмы-производители двуствольных дробовиков стран ПМК маркируют на стволах их массу, однако эта маркировка совсем не упоминается в перечне обязательных. Смысл ее нанесения совершенно неясен, на ружьях российского производства не встречается.
Смысл ее нанесения совершенно неясен, на ружьях российского производства не встречается.
4.4. Знаки собственников. По особым указаниям на стволах отдельных партий ружей наносились надписи с названиями фирм-импортеров. Например, на одной из партий ружей ИЖ-81 справа на стволе по просьбе заказчика было написано лазером:
IMPORTED BY CAI ST.ALB.VT
please read owners manual before firing
Последняя строка переводится как «перед использованием, пожалуйста, читайте руководство по эксплуатации». Такая надпись сейчас обычна не только для американских охотничьих ружей.
Иногда по частным заказам на заводе (и не только) на личинке или спусковой скобе ружья наносились причудливые монограммы владельца. Монограмма или дарственная надпись может быть нанесена и на специальной пластине, врезанной в приклад.
Вот и всё, что вкратце можно сказать о надписях на ижевских ружьях.
В.Вальнев, инженер-оружейник
Читать киберфантастику онлайн на Призрачных мирах
Патроны и калибры для ИЖ 27 (МР 27)
Наиболее популярный калибр ИЖ-27 – 12 × 76 (под диаметр ствола 18,4 мм). Также существует 20-й калибр (также под гильзу 76 мм) и 16-й (под 70 мм гильзу). Усиленная версия с диаметром ствола в 10,3 мм под .410 калибр – довольно редкая и в настоящее время ее можно встретить в основном у заядлых охотников и у коллекционеров.
Также существует 20-й калибр (также под гильзу 76 мм) и 16-й (под 70 мм гильзу). Усиленная версия с диаметром ствола в 10,3 мм под .410 калибр – довольно редкая и в настоящее время ее можно встретить в основном у заядлых охотников и у коллекционеров.
МР-27 – достаточно неприхотливое оружие и подходит под любую дробь. А вот с пулевыми боеприпасами не все так однозначно. Например, для чока и получока, цилиндра 0 и 0,25 мм лучше использовать подкалиберные пули, которые меньше калиберных и помещены в пластиковый контейнер. За счет этого боеприпас пролетает по каналу ствола без помех. При прохождении через дульное сужение, пластик сдавливается, не нанося при этом повреждений стволу. Если же мы говорим о калиберных пулях, диаметр которых превышает размер сужения, происходит следующая ситуация: из-за несоответствия размеров перед дульным сужением в стволе возникает сильное напряжение. Со временем это приводит к разрыву ствола. Поэтому, от использования калиберных пуль стоит отказаться.
Карабин ИЖ-27 СССР
Как выбрать ружье МР 27: правильное поведение в магазине
Если вы твердо решили купить вертикальную двустволку ИЖ-27, определились с модификацией и исполнением, осталось только посетить оружейный магазин и сделать покупку. На что же обратить внимание в магазине?
На что же обратить внимание в магазине?
Длина приклада
Оружие нужно «примерить», точно так же как мы примеряем перед покупкой одежду или обувь. Половина удачи на охоте зависит именно от того, насколько удобной для вас будет гладкостволка. Иногда охотники приходят в магазин за одним ружьем, а покупают совсем другое – то, которое идеально легло. Когда, держа ружье наготове, ненадолго закрываешь глаза, а при открытии видишь, что мушка осталась на прежнем месте.
И несколько практических советов:
- 1. Проводите примерку прикладистости в той одежде, в которой в дальнейшем будете ездить на охоту (или заниматься спортивной стрельбой).
- 2. В ИЖ 27 приклады можно менять. Попробуйте подобрать несколько подходящих вариантов, например, для охоты в летнее и зимнее время года, в межсезонье.
Прицельная планка
Перед покупкой убедитесь в качестве сварных швов и ровности планки прицела. Для контроля качества пайки прозвоните планку донцем гильзы или небольшим молотком. Внимательно прослушайте, какой звук при этом издается. Он должен быть ровным и без проседаний. Не забудьте убедиться и в надежности межствольной пайки. Осмотрите стыки, убедитесь в отсутствии трещин, пор и сколов.
Внимательно прослушайте, какой звук при этом издается. Он должен быть ровным и без проседаний. Не забудьте убедиться и в надежности межствольной пайки. Осмотрите стыки, убедитесь в отсутствии трещин, пор и сколов.
Работа эжектора
Исправность автоматического выбрасывателя стреляных гильз проверить легко: когда после выстрела при раскрытии стволов слышно щелчок – это работает эжектор, выпихивает отстрелянные гильзы, тем самым обеспечивая скорость перезарядки. Ведь если животное позволят сделать несколько выстрелов, можно, переломив ружье, сразу его перезарядить.
Стволы
Состоянию стволов также следует уделить внимание перед покупкой. Убедитесь при тщательном визуальном осмотре в отсутствии вздутий, раковин и прочих дефектов. Должное внимание уделите геометрическим параметрам. Прежде всего, стволы должны быть ровными.
Ровность легко проверить несколькими способами:
- 1. Вложите гильзу без капсюля в патронник.
- 2. Наведите патронник с гильзой на источник света.

- 3. Осмотр следует проводить со стороны дул.
- 4. Кольца должны быть круглыми, четкими, ровными. Их кривизна или смещение укажет на брак.
Ружье ИЖ-27Е
[править] Односпусковой механизм
Ружье Иж 27Е-1С
является первой моделью односпускового охотничьего ружья, освоенной отечественной промышленностью в крупносерийном производстве. Сам же механизм с одним спусковым крючком был разработан в 1965 году Е.Губиным и впервые применен ижевскими мастерами в спортивно-тренировочном ружье Иж 25. Механизм отличается простотой устройства и высокой надежностью в работе.
Преимущества односпусковых механизмов в двуствольных ружьях очевидны, и поэтому они все шире применяются не только в спортивных, но и в охотничьих ружьях (особенно в США). Односпусковой механизм позволяет в более короткий промежуток времени произвести два выстрела последовательными нажатиями на спусковой крючок, без отвлечения внимания на второй спусковой крючок. Последнее способствует сохранению устойчивости ружья во время стрельбы, позволяет в полной мере использовать преимущества ложи с пистолетной шейкой, на которой кисть руки занимает наиболее удобное и естественное положение.
Последнее способствует сохранению устойчивости ружья во время стрельбы, позволяет в полной мере использовать преимущества ложи с пистолетной шейкой, на которой кисть руки занимает наиболее удобное и естественное положение.
Комплектующие
Опытные охотники знают, что помимо самого оружия и патронов, дульных сужений, потребуется приобрести еще ряд дополнительных комплектующих и аксессуаров. Рассмотрим их.
Сменный приклад ИЖ 27 (МР 27)
Как уже отмечалось, приклады на ИЖ-27 выпускаются самые разные: из дерева, пластика, различной длины.
Затыльники
Каждый стрелок имеет индивидуальные анатомические особенности. И подобрать подходящий по размеру затыльник, с идеальной посадкой, довольно сложно. Но не в случае с ИЖ-27! предлагает по-настоящему королевский выбор затыльников из дерева и полимеров. Классический затыльник из дерева способствует быстрой вскидке оружия, резиновый – обладает хорошей амортизацией и эффективно смягчает удары на плечо, имеет превосходную морозостойкость. Резиновые затыльники наиболее распространены и выпускаются под разные длины прикладов. Затыльники из текстолита имеют шероховатую поверхность. Эта особенность дает возможность надежно удерживать «гладкостволку» за счет того, что ружье не скользит по одежде. Текстолит – прочный и морозостойкий материал.
Резиновые затыльники наиболее распространены и выпускаются под разные длины прикладов. Затыльники из текстолита имеют шероховатую поверхность. Эта особенность дает возможность надежно удерживать «гладкостволку» за счет того, что ружье не скользит по одежде. Текстолит – прочный и морозостойкий материал.
В точности как приклад, затыльник нужно «примерить». Он не должен мешать вскидке ружья. Некоторые опытные охотники имеют несколько затыльников на разные сезоны: например, для лета, когда на охотнике нет толстого бушлата, подбираются затыльники большей толщины.
Мушки и прицелы
Мушка на ИЖ 27 в стандартной комплектации идет толстая. Для точного прицеливания рекомендуется ее сразу же сменить на более тонкие варианты, которые завод предлагает в огромном количестве: стальные, удлиненные, оптоволоконные и светонакопительные.
В нашем оружейном ломбарде вы можете приобрести гладкоствольное оружие бу, как иж 27, так и других марок.
Наши контакты: [email protected]
Также обратите внимание на наш Telegram канал и то, что мы доступны для связи в WhatsApp по номеру:
+7
Аналоги
- Ближайшим конкурентом на российском рынке для МР-27М является вторая «народная» винтовка – ТОЗ-34.
 Оружие имеет свои достоинства и недостатки по сравнению друг с другом, так что этот спор не утихает и сегодня.
Оружие имеет свои достоинства и недостатки по сравнению друг с другом, так что этот спор не утихает и сегодня. - Из зарубежных можно назвать двустволки «Меркель-2000» и «Меркель-2001». Выпускаются ружья в 12, 20 и 28 калибрах. С 2002 года на них установлен патронник длиной в 76 мм. Стоимость немецких винтовок, конечно, заметно выше.
МР-27М – простая и надежная вертикальная двустволка. Производится линейка более 40 лет, но пользуется непреходящей популярностью благодаря точности, прикладистости и доступной стоимости.
Отзыв владельца на ружье МР-27М дан в этом видео:
ИЖ-27 — Вики
Материал из Википедии — свободной энциклопедии
Перейти к навигацииПерейти к поиску
У этого термина существуют и другие значения, см. Иж.
ИЖ-27 — советское двуствольное охотничье ружьё, предназначенное для промысловой и любительской охоты[2].
Содержание
- 1 История
- 2 Описание
- 3 Варианты и модификации
- 4 Примечания
- 5 Литература
- 6 Ссылки
История
ИЖ-27 было разработано в начале 1970х годов[4] под руководством А. А. Климова. В 1972 году демонстрационный образец ружья ИЖ-27 был официально представлен на выставке охотничьего оружия, боеприпасов и снаряжения в Иркутске (проходившей в здании Иркутского сельскохозяйственного института)[5]. Серийно выпускается с 1973 года[1].
А. Климова. В 1972 году демонстрационный образец ружья ИЖ-27 был официально представлен на выставке охотничьего оружия, боеприпасов и снаряжения в Иркутске (проходившей в здании Иркутского сельскохозяйственного института)[5]. Серийно выпускается с 1973 года[1].
Является результатом глубокой модернизации ружья ИЖ-12. В сравнении с ИЖ-12, у ИЖ-27 была изменена форма ложи и цевья, введена вентилируемая прицельная планка и изменена форма коробки в местах соединения с ложей[6].
По состоянию на начало 1985 года ружья ИЖ-27 и ТОЗ-34 являлись самыми распространёнными моделями охотничьих ружей в СССР[7], также они предлагались на экспорт в другие страны[8].
18-24 января 1990 года на выставке «КОНСУМЭКСПО-1990» в Москве была представлена версия ружья ИЖ-27 со сменными дульными насадками (предложенными также для ИЖ-18М и ИЖ-43)[9].
Только в период до 1 января 1998 года было выпущено 575 701 шт. ружей ИЖ-27 (и их модификаций), 560 197 шт. ружей ИЖ-27Е (и их модификаций), 22 004 шт. ружей ИЖ-27ММ и ИЖ-27М-1С-М, а также 11 139 шт. ружей ИЖ-27ЕММ и ИЖ-27ЕМ-1С-М[10].
ружей ИЖ-27Е (и их модификаций), 22 004 шт. ружей ИЖ-27ММ и ИЖ-27М-1С-М, а также 11 139 шт. ружей ИЖ-27ЕММ и ИЖ-27ЕМ-1С-М[10].
С 1999 года начался импорт ружей ИЖ-27 в США через компанию «European American Armory»[11]. Ещё некоторое количество ружей ИЖ-27 в 2005 — 2009 годы было выпущено по заказу американской оружейной компании «Remington Arms», они продавались в США под наименованием Remington Spartan 310[12].
До сентября 2008 года ружьё производилось под наименованием ИЖ-27[3].
Выпущено более 1,5 миллиона ружей ИЖ-27 всех модификаций[3] — таким образом, это ружьё стало самым распространённым охотничьим ружьём в России[13].
Описание
Стволы ружья изготовлены из ствольной стали 50РА (и для повышения прочности термообработаны по всей длине), отъёмные, расположены в вертикальной плоскости. Стволы запрессованы в муфту и соединены между собой пайкой на боковых планках[2].
- дульное сужение нижнего ствола — 0,5 мм (получок)[2]
- дульное сужение верхнего ствола — 1 мм (полный чок)[2]
Ружьё простое в эксплуатации и обслуживании — для доступа к ударно-спусковому механизму необходимо отвинтить продольный ложевой болт и отделить цевье[2].
Приклад и цевьё изготовлены из дерева. В зависимости от варианта исполнения, ложа может быть прямая или полупистолетная, приклад может быть оборудован пластмассовым затыльником или резиновым амортизатором[2].
Варианты и модификации
Ружьё выпускалось в серийном, в том числе экспортном, штучном и сувенирном исполнении в нескольких различных модификациях[6]:
- ИЖ-27 — первая модель, серийный выпуск начат в 1973 году. Стандартные серийные ружья выпускались со стволами 730 мм, но по специальным заказам изготавливали штучные ружья со стволами длиной 680, 700, 750 и 760 мм[2]
- ИЖ-27-1С — вариант ИЖ-27 с одним спусковым крючком, который обеспечивал поочерёдную последовательность стрельбы («нижний ствол, затем верхний ствол»).
 Спусковой механизм разработал в 1965 году мастер-оружейник Е. И. Губин и изначально он устанавливался на спортивное ружье ИЖ-26[14]. Серийное производство началось в середине 1970-х годов[6].
Спусковой механизм разработал в 1965 году мастер-оружейник Е. И. Губин и изначально он устанавливался на спортивное ружье ИЖ-26[14]. Серийное производство началось в середине 1970-х годов[6]. - ИЖ-27Е — ружьё с эжектором конструкции Г. Я. Протопопова, серийный выпуск начат в 1973 году[2]
- ИЖ-27Е-1С — спортивно-охотничье ружьё с эжектором (который может быть отключён при необходимости), с одним спусковым крючком, который обеспечивал поочерёдную последовательность стрельбы («нижний ствол, затем верхний ствол»). Серийное производство началось в середине 1970-х годов[6], разработано при участии конструкторов Э. А. Корепанова и А. В. Головина, в 1976 году на Международной выставке в Брно (ЧССР) ружьё было награждено золотой медалью[15].
- ИЖ-27СТ — спортивное ружьё для стрельбы на траншейном стенде, с 760 мм стволами[16]
- ИЖ-27СK — спортивное ружьё для стрельбы на круглом стенде, с 660 мм стволами[16]
- МР-27М (ранее называлось ИЖ-27М) — ружьё без эжектора, с двумя спусковыми крючками.

- ИЖ-27M «Юниор» — облегченный вариант ИЖ-27М для женщин и подростков, с уменьшенной длиной приклада
- ИЖ-27ММ — вариант ИЖ-27М под патрон 12×76 мм «Магнум»
- МР-27ЕМ (ранее называлось ИЖ-27ЕМ) — ружьё с эжектором селективного типа — выбрасывает только стреляную гильзу и, при необходимости, может быть отключён. Имеет два спусковых крючка.
- МР-27М-1С (ранее называлось ИЖ-27М-1С) — ружьё без эжектора. Ударно-спусковой механизм с одним спусковым крючком позволяет вести стрельбу из обоих стволов в нужной последовательности. Последовательность стрельбы меняется путём нажатия на спусковой крючок сзади.
- МР-27ЕМ-1С (ранее называлось ИЖ-27ЕМ-1С) — ружьё с эжектором селективного типа — выбрасывает только стреляную гильзу и, при необходимости, может быть отключён. Ударно-спусковой механизм с одним спусковым крючком позволяет вести стрельбу из обоих стволов в нужной последовательности.
 Последовательность стрельбы меняется путём нажатия на спусковой крючок сзади.
Последовательность стрельбы меняется путём нажатия на спусковой крючок сзади.
Ружьё ИЖ-27 послужило базой для создания спортивных ружей ИЖ-25 и ИЖ-39 для круглого и траншейного стендов[1] (сейчас уже не выпускаются)[17], а также для комбинированных ружей MP-94[18].
Примечания
- ↑ 1 2 3 Описание ИЖ-27 на сайте www.extreme.by (неопр.). Дата обращения: 22 февраля 2011. Архивировано 22 декабря 2011 года.
- ↑ 1 2 3 4 5 6 7 8 9 10 В. Владимиров. Ружье ИЖ-27 // журнал «Охота и охотничье хозяйство», № 2, 1973.
 стр.30
стр.30 - ↑ 1 2 3 4 МР-27М — АО «ИЖЕВСКИЙ МЕХАНИЧЕСКИЙ ЗАВОД» (неопр.) (недоступная ссылка). Дата обращения: 26 октября 2017. Архивировано 7 октября 2017 года.
- ↑ «Двуствольное охотничье внутрикурковое ружьё 12 калибра с вертикально расположенными стволами ИЖ-27 … создано на базе находящегося в производстве ружья ИЖ-12 с целью его замены«
Г. Карагодин, Н. Аксенов. Тенденции развития охотничьего оружия // журнал «Охота и охотничье хозяйство», № 4, 1970. стр.28-29 - ↑ Э. Штейнгольд. Выставка-продажа охотничьего оружия // журнал «Охота и охотничье хозяйство», № 11, 1972. стр.32-33
- ↑ 1 2 3 4 А. В. Кузьминский. Оружие для охотника: практическое пособие / под общ.
 ред. А. Е. Тараса М., ООО «Издательство АСТ», 2002. стр.176-177
ред. А. Е. Тараса М., ООО «Издательство АСТ», 2002. стр.176-177 - ↑ Е. В. Кудрявцев. Охота (краткий справочник). 3-е изд., доп. М., Агропромиздат, 1985. стр.15
- ↑ Ружьё двуствольное ИЖ-27 // Охотничье и спортивное оружие. М., Внешторгиздат. 1989.
- ↑ Н. Аксенов. «КОНСУМЭКСПО-1990»: оружейные новинки // журнал «Охота и охотничье хозяйство», № 6, 1990. стр.28-29
- ↑ Михаил Драгунов. Анатолий Климов. Неизвестный конструктор известных ружей. К 80-летию со дня рождения // журнал «Калашников. Оружие. Боеприпасы. Снаряжение», № 1, 2001. стр.4-6
- ↑ EAA Baikal IZH-27 // J. B. Wood. The Gun Digest Book of Firearms Assembly / Disassembly. Part V : Shotguns. 2nd edition. Krause Publications, 2002. pages 101-109
- ↑ Baikal IZH-27 / Remington SPR-310 // Michael E. Haskew. Rifles & Muskets from 1450 to present day. London, Amber Book Ltd., 2015
- ↑ Описание ИЖ-27 на сайте www.safari-club.ru (неопр.
 ). Дата обращения: 22 февраля 2011. Архивировано 11 января 2012 года.
). Дата обращения: 22 февраля 2011. Архивировано 11 января 2012 года. - ↑ Н. Изметинский, Л. Михайлов. ИЖ-27Е-27-1С — охотничье ружье с одним спуском // журнал «Охота и охотничье хозяйство», № 9, 1978. стр.30-32
- ↑ Ижевские охотничьи ружья // журнал «Охота и охотничье хозяйство», № 8, 1977
- ↑ 1 2 А. В. Сицко, М. М. Блюм, И. Б. Шишкин. Советы начинающему охотнику. М., «Агропромиздат», 1991. стр.227
- ↑ Перечень выпускаемого оружия на сайте производителя (неопр.) (недоступная ссылка). Дата обращения: 23 февраля 2011. Архивировано 1 мая 2011 года.
- ↑ «Конструктивно МР-94 является усовершенствованной версией безотказной двуствольной вертикалки МР-27»
Галина Валеева. Комбинированные ружья MP-94 // журнал «Калашников. Оружие. Боеприпасы. Снаряжение», № 7, 2015. стр.52-55
стр.52-55
Литература
- Л. Е. Михайлов, Н. Л. Изметинский. Ижевские охотничьи ружья. 2-е изд., испр. и доп. Ижевск, изд-во «Удмуртия», 1982. стр.160-187
- Э. Корепанов. Один спуск на два ствола // журнал «Охота и охотничье хозяйство», № 11-12, 1992. стр.22-23 [о спусковом механизме ИЖ-27-1С]
- Виктор Гордиенко. Главный калибр российского охотника // журнал «Мастер-ружьё», № 1 (50), 2001. стр.112-117
- В. Вальнев. Замена бойка на ружье ИЖ-27 // журнал «Охота и охотничье хозяйство», № 7, 2007. стр.30-31
- В. Вальнев. ИЖ-27: устройство и ремонт предохранительного механизма // журнал «Охота и охотничье хозяйство», № 1, 2010. стр.32-33
- Валеева Г. ИЖ-27 вчера и МР-27 сегодня. // Журнал «Калашников». — 2012. — № 8. — С. 40—45.
- Universal/Baikal IJ-27E shotgun // журнал «American Rifleman», December 1975
- Baikal IZH-27 // журнал «American Rifleman», May 2001
Ссылки
Содержимое этой статьи нуждается в чистке. Текст содержит много маловажных, неэнциклопедичных или устаревших подробностей. Пожалуйста, улучшите статью в соответствии с правилами написания статей. |

- http://www.gun.ru/oxota1003.htm Архивная копия от 30 декабря 2010 на Wayback Machine
- http://www.tropaohotnika.ru/izh-27.php Архивная копия от 24 мая 2011 на Wayback Machine
- http://www.ohoter.ru/464-oxotniche-ruzhe-izh-27.html Архивная копия от 18 марта 2011 на Wayback Machine
- http://www.strelok-kmv.ru/izh-27m.html Архивная копия от 12 июня 2010 на Wayback Machine
- http://bestoxotnik.ru/izh-27/ Архивная копия от 19 марта 2013 на Wayback Machine
- Виктор Вальнев — Устройство и ремонт предохранительного механизма ружья Иж-27. Архивная копия от 28 января 2020 на Wayback Machine Авторский сайт «Переулок оружейников»
- Ружьё ИЖ-27, описание, паспорт: http://huntset.com/page83.html Архивная копия от 28 октября 2015 на Wayback Machine
- IZh-27 Архивная копия от 12 января 2019 на Wayback Machine / Internet Movie Firearms Database (использование в кинофильмах)
основные технические характеристики, модификации, правила хранения и ухода
Двуствольное ружье ИЖ-27 является модификацией предыдущей модели — ИЖ-12. Инженеры Ижевского механического завода внесли довольно существенные изменения в выпускавшийся прежде вариант. В результате эта вертикалка стала самым продаваемым охотничьим оружием в СССР и России. Чтобы разобраться в причинах популярности ружья, необходимо изучить его технические характеристики и устройство.
Инженеры Ижевского механического завода внесли довольно существенные изменения в выпускавшийся прежде вариант. В результате эта вертикалка стала самым продаваемым охотничьим оружием в СССР и России. Чтобы разобраться в причинах популярности ружья, необходимо изучить его технические характеристики и устройство.
История создания и модификации
В конце 60-х годов на Ижевском механическом заводе (ИМЗ) было пряно решение о модернизации гладкоствольной модели ИЖ-12. Это было связано с наличием у выпускавшегося тогда ружья нескольких существенных недостатков. Создание нового оружия было доверено группе специалистов, которыми руководил Климов А. А. В результате была представлена модель ИЖ-27, в современной классификации — ружьё МР 27. Серийный выпуск двустволки стартовал в 1973 году.
Неизменным осталось расположение стволов в вертикальной плоскости. Первыми были модифицированы цевье и ложе. Конструкторы постарались сделать эти элементы конструкции максимально удобными и эргономичными.
Прицельную планку оснастили вентиляционными отверстиями, что сделало более удобным прицеливание. Особенно это заметно в жаркую пору года, благодаря эффективному охлаждению. Также вместо ручного предохранителя был установлен автоматический.
Конструкторы решили установить и устройство для перехватывания курков (интерсепторы). Кроме этого, впервые за всю историю предприятия, выпускаемое им ружье было оснащено эжектором для гильз. Чтобы оружие было более удобным в эксплуатации и обслуживании, модификации подверглось место соединения коробки с ложей.
Благодаря резиновому затыльнику на прикладе, винтовка стала еще более удобной в применении.
На сегодняшний день завод выпускает несколько модификаций вертикалки:
- ИЖ 27 М. Характеристики ружья практически аналогичны базовой модели, но отсутствует эжектор.
- 27М-1С. Вертикалка имеет только один курок. Для смены последовательности выстрелов спусковой курок необходимо слегка подать вперед до появления щелчка.
 Это весьма удобно для фиксации действий, но если охота проходит в тихую погоду, зверь может услышать звук
Это весьма удобно для фиксации действий, но если охота проходит в тихую погоду, зверь может услышать звук - ИЖ 27ЕМ. В этой модификации эжектор предназначен для выброса только отстрелянной гильзы.
- ИЖ 27Е-1С. Имеет один спусковой курок и эжектор.
Технические характеристики и устройство
На сегодняшний день было выпущено более 1,6 млн единиц этого ружья. Кроме серийных моделей, встречаются и сувенирные изделия. Среди основных технических характеристик стоит отметить:
- Калибр. Наиболее популярными являются модели 12 и 16 калибра, но также выпускаются экземпляры 20, 28, 32 и 410.
- Длина патронника. Этот размер составляет 70 и 76,2 мм.
- Дульные сужения. Для нижнего ствола показатель равен 0,5 м, а для верхнего — 1 мм.

- Длина ружья ИЖ 27 в сборе составляет 720—730 мм.
- Вес. В зависимости от модели, может равняться от 3,1 до 3,6 кг.
Стволы оснащены вентилируемой прицельной планкой. Патронники, а также канал ствола хромированы. Все наружные поверхности ружья прошли процедуру оксидирования либо анодирования.
Ударно-спусковой механизм смонтирован в коробке и частично на расположенном отдельно нижнем основании. Кроме этого, УСМ оснащен внутренними курками. Механизм вертикалки обладает интерсепторами, предназначенными для перехвата курков в случае неожиданного падения. Это сделало ружье более безопасным. Боевые пружины витые и имеют цилиндрическую форму. Также в конструкции ударно-спускового механизма есть инерционный разобщитель. В результате предотвращается возможность двойного выстрела.
В период с 1972 по 1977 год все ружья выпускались с двойным курком. Однако затем была создана модификация с одним крючком. Изначально она создавалась для спортивного оружия модели ИЖ-25. Когда ружье заряжено, очередность выстрелов является стандартной — верхний-нижний ствол.
Когда ружье заряжено, очередность выстрелов является стандартной — верхний-нижний ствол.
Для изменения очередности необходимо оттянуть курок вперед до появления щелчка. Следует помнить, что после новой перезарядки автоматически восстановится прежняя очередность. Бойки обладают довольно большим ходом — до 2,2 мм, а также оснащены возвратными пружинами. У любителей охоты никаких претензий к работе УСМ за все время выпуска модели не возникало.
Приклад на серийных изделиях изготовлен из бука либо ореха и имеет шейку пистолетного типа. В штучных моделях, изготовленных по индивидуальному заказу, этот элемент конструкции может иметь любую форму.
Сравнение с ружьем ТОЗ-34
Эти модели являются основными конкурентами. В ТОЗ-34 и ИЖ-27 стволы расположены в вертикальной плоскости, обладают примерно одинаковыми характеристиками и стоимостью. Также следует заметить, что обе модели пользуются высокой популярностью у отечественных охотников. Однако между ними есть несколько существенных различий.
Однако между ними есть несколько существенных различий.
Наиболее существенным среди них является качество стволов. Принято считать, что оно несколько лучше у ТОЗ-34. Можно отметить и другие важные различия:
- Боевые пружины ружья Ижевского завода — спиральные, и поэтому более надежные. В ТОЗ-34 эти элементы конструкции фигурные, что снижает срок их использования.
- На ижевской вертикалке установлено съемное цевье. При его частом демонтаже возможно появление трещин на древесине. У конкурента цевье несъемное и этого недостатка лишено.
- Одним из самых серьезных недостатков ТОЗ-34 является рычаг взвода ударников. Этот элемент конструкции имеет не самое высокое качество изготовления, что приводит к частым поломкам.
- Вес ТОЗ-34 меньше, что можно считать не только достоинством, но и недостатком. Это зависит в первую очередь от выбранного стиля охоты.
- Запорные механизмы у каждой модели имеют хорошее качество и могут исправно работать длительное время.

Обе модели имеют ряд преимуществ и недостатков. При их выборе следует в первую очередь ориентироваться на личные предпочтения.
Рекомендации по выбору
Не имеет принципиального значения, где приобретается ружье. Даже если вертикалка новая, есть несколько нюансов, о которых стоит знать, чтобы остаться довольным покупкой. Среди них можно выделить следующие:
- Необходимо обратить внимание на качество соединения стволов. Планка должна быть хорошо пропаяна. Никакого зазора либо белого налета между ней и стволом быть не должно.
- Не допускается овальность дульных срезов, а толщина стенки ствола должна быть одинаковой.
- Обязательно проводится осмотр внутренней поверхности ствольного канала на предмет отсутствия просветов и пятен.
- Не допускается наличие изгибов, изломов и выпуклостей на прицельной планке. Этот элемент конструкции должен быть идеально ровным.
- Оружие должно легко разбираться.

- Необходимо проверить деревянные поверхности вертикалки на наличие трещин и сколов.
Если ружье потряхивать в собранном виде, то никакого дребезжания механизмов быть не должно.
Уход и хранение
Ружье необходимо хранить в разобранном состоянии в сухом месте. Также необходимо предварительно спустить курки, чтобы предотвратить повышенный износ боевых пружин. Все детали в обязательном порядке должны быть смазаны оружейным маслом, это позволит предотвратить возможность их коррозии. Если вертикалка не будет использоваться длительное время, то все металлические элементы конструкции следует покрыть смазкой густой консистенции.
После каждой стрельбы оружие предстоит тщательно вычистить и смазать. Хромирование ствольных каналов чаще всего является слабым местом модели. Только регулярный грамотный уход позволит избежать появления ржавчины. Если на каналах ствола появились признаки оцинковки, то их необходимо быстро удалить с помощью мягкого ершика, изготовленного из меди. Также можно использовать специальные чистящие средства, купить которые можно в специализированных магазинах.
Также можно использовать специальные чистящие средства, купить которые можно в специализированных магазинах.
При появлении шата стволов следует проверить степень натяжения специального винта. Для решения поставленной задачи предстоит выкрутить два шурупа, после чего проверить стяжной винт. Не рекомендуется щелкать спусковыми курками вхолостую. Если в стволах отсутствуют патроны, то бойки и посадочные гнезда могут деформироваться. Для тренировки дома следует использовать стреляные гильзы либо приобрести специальные флеш-патроны.
Ружье ИЖ-27 отлично подходит не только для любителей охоты, но и для промысловиков. Это проверенное временем оружие, которое активно используется миллионами охотников. Благодаря привлекательной стоимости и хорошему качеству, конкурентов у этой вертикалки практически нет.
- Автор: Владимир
- Распечатать
Оцените статью:
(0 голосов, среднее: 0 из 5)
Поделитесь с друзьями!
история создания и конструктивные особенности ружья
ИЖ-27 12 калибра (он же MP-27) — самое популярное российское ружье среди моделей с вертикальным расположением стволов. Оружие отличается высокими тактико-техническими характеристиками, приятной эргономикой, уникальными внешними данными и возможностями для установки апгрейда. Благодаря этому ружье 12 или 16 калибра известно не только среди охотников, но и профессиональных спортсменов, а также простых ценителей гладкоствольных ружей.
Оружие отличается высокими тактико-техническими характеристиками, приятной эргономикой, уникальными внешними данными и возможностями для установки апгрейда. Благодаря этому ружье 12 или 16 калибра известно не только среди охотников, но и профессиональных спортсменов, а также простых ценителей гладкоствольных ружей.
Содержание:
- История создания
- Конструкция
- Описание и технические характеристики
- Модификации ИЖ-27 (МР-27)
- Список штучных моделей
 org/ListItem»> Достоинства и недостатки
org/ListItem»> Достоинства и недостатки- Как выбрать: новое или БУ
- Хранение и уход
- Тюнинг
- Отзывы охотников
История создания
История создания ИЖ-27 берет свое начало в 1970 году. В то время перед инженерами Ижевского механического завода встала важная задача. Необходимо было создать универсальное гладкоствольное ружье, которое бы использовалось охотниками и спортсменами в течение нескольких десятилетий. В качестве образца было принято решение взять ИЖ-12 — долгое время именно эта двустволка с вертикальным расположением стволов считалась самой популярной. За устранение недостатков и модернизацию существующих систем взялся инженер по фамилии Климов.
Разработка ИЖ-27 продолжалась в течение почти трех лет, так что первое ружье сошло с конвейера в 1973 году. Новая модель сразу же произвела огромное впечатление на простой люд. Первая партия в 10 тысяч ружей была раскуплена буквально за несколько дней. Ружье покупалось охотниками и спортсменами. Первые выбирали в основном 12 калибр стволов, вторые — останавливали свой выбор на 16-ом. За все время продаж было реализовано более полумиллиона копий легендарного ружья.
В дальнейшие годы ИЖМАШ занимался разработкой модификации для столь удачного проекта. Одними из первых улучшенных ружей стали ИЖ-27М (модифицированное) и ИЖ-27Е (эжекторное). Также ижевский механический завод регулярно получал просьбы разработать несколько штучных модификаций, отличающихся более дорогим внешним видом. Выпущенные модели ИЖ-27-Ш1 и ИЖ-27-Ш2 отличались от своих старших собратьев наличием гравировки на металле, прикладом из дорогих пород дерева, а также серебряными и золотыми вставками на ложе.
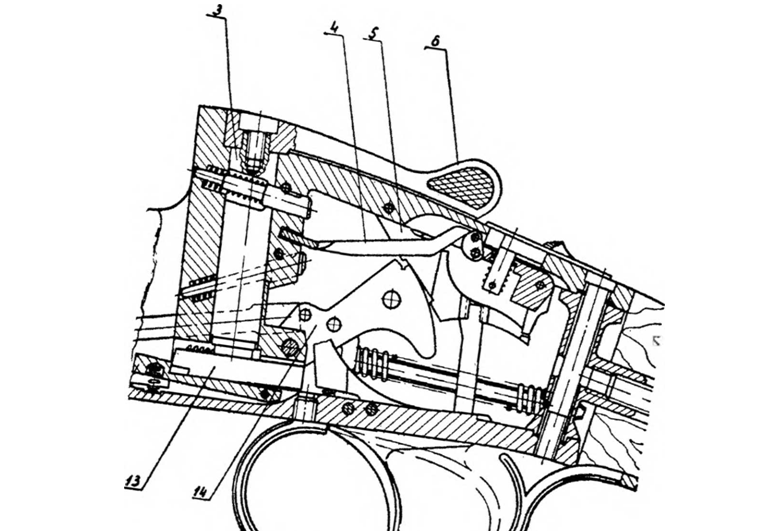
Успех ИЖ-27 оказался настолько большим, что моделью заинтересовалась американская оружейная компания Remington Arms. ИЖМЕХУ было предложено продать лицензию на производство оружия, на что компания согласилась после нескольких месяцев раздумий. Американский ИЖ-27 стал называться Remington Spartan 310, однако популярности в Штатах ружье не получило — видимо, штамп «Произведено в России» вызывает у мировых охотников куда больше доверия. И по сей день многие любители промысла из-за границы продолжают покупать легендарный ИЖ-27 и его модификации.
Классическое ружье иж-27 в разобранном видек содержанию ↑
Конструкция
Чтобы понимать, что представляет собой то или иное оружие, необходимо как следует изучить его основные конструктивные особенности. Ружье ИЖ-27 — это классическая двустволка с вертикальным расположением стволов, которая выпускается в 12, 16, 20 и других калибрах. От всем известного прототипа ИЖ-12 модель отличается не сильно, однако инженеры Ижевского механического завода не могли не внести несколько изменений:
- Более эргономичная и удобная форма цевья и ложе.
 На деревянной части стал присутствовать вырез под щеку, а пистолетная рукоять обрела оптимальную форму, хорошо подходящую под руку не только взрослого мужчины, но и девушки.
На деревянной части стал присутствовать вырез под щеку, а пистолетная рукоять обрела оптимальную форму, хорошо подходящую под руку не только взрослого мужчины, но и девушки. - Наличие вентиляционных отверстий, расположенных на прицельной планке. Простая, но поистине гениальная задумка ИЖМЕХА в дальнейшем стала использоваться во многих образцах гладкоствольных вертикалок, имеющих большую парусность.
- Замена ручного предохранителя на автоматический. Теперь владельцу не требовалось самостоятельно устанавливать оружие в безопасный режим после взведения стволов. Спусковой крючок блокировался в тот момент, когда происходило переламывание ружья.
- Оружие обзавелось интерцепторами — специальными курками, которые выполняют функцию запирателя стволов. Такая система присутствовала и в других моделях ружей от ИЖМЕХА, однако именно здесь была доведена до совершенства.
- Наличие встроенного эжектора для гильз. Система служит для одной цели — выбрасывания использованных патронов из ствола в момент открытия патронника.
 Механизм оказался просто незаменим при использовании модели в сильный мороз.
Механизм оказался просто незаменим при использовании модели в сильный мороз. - Тыльная часть приклада обзавелась резиновой насадкой, служащей для повышения уровня эргономики. Большую часть отдачи от выстрела резина забирала на себя, так что использовать ружья стало гораздо приятнее.
- Качественное соединение ствольной коробки с ложем и прикладом. Многие сравнивают эту технологию с той, что существовала во времена фараонов. Человек не сможет просунуть между плитами пирамиды даже иголку, точно также как и между деревом и металлом в ИЖ-27.
Во всем остальном конструкция осталась схожа с моделью ИЖ-12. К примеру, для производства металлических деталей продолжала использоваться сталь А50, отличающаяся повышенным содержанием углерода, а также прошедшая процедуру холодной ковки. В дальнейшем конструкция модели претерпела множество изменений, однако более подробно об этом мы еще поговорим в разделе «Модификации».
к содержанию ↑
Описание и технические характеристики
Ружье ИЖ-27 пользуется такой популярностью не только потому, что превзошло свой прототип. Модель также отличается непревзойденными боевыми характеристиками — главным фактором, который ценят любители охотничьего промысла. Найти эти показатели можно в оружейном паспорте, который всегда идет в комплекте с ружьем. Однако некоторые значения могут отличаться (зависит от калибра). Так что информация из следующей таблицы будет актуальна лишь для ИЖ-27 12 калибра:
Модель также отличается непревзойденными боевыми характеристиками — главным фактором, который ценят любители охотничьего промысла. Найти эти показатели можно в оружейном паспорте, который всегда идет в комплекте с ружьем. Однако некоторые значения могут отличаться (зависит от калибра). Так что информация из следующей таблицы будет актуальна лишь для ИЖ-27 12 калибра:
| Характеристика | Параметр |
|---|---|
| Длина ствола | 750 мм |
| Используемый патрон | 18,6 мм |
| Длина патронника | 76 мм |
| Чок (сужение) | 1 мм |
| Вес оружия | 3,7 кг |
Мощность выстрела составляет 75 Дж, а скорость баллистического снаряда — 365 м/с. Таких показателей вполне хватает, чтобы охотиться на взрослого кабана или даже медведя средних размеров (при условии согласованного патрона). Также охотникам понравится тот факт, что ружье может быть применимо на дистанции до 100 метров — кучность боя сохраняет показатель от 55% до 60%. Именно поэтому многие заряжают ружье пулей и отправляются за добычей хищника, ведь опасного зверя можно держать на расстоянии.
Именно поэтому многие заряжают ружье пулей и отправляются за добычей хищника, ведь опасного зверя можно держать на расстоянии.
Несмотря на высокую мощность, срок жизни оружия составляет от 5 до 10 лет. А при условии качественного обслуживания показатель можно и вовсе увеличить в разы. Все это благодаря наличию хромированного покрытия внутри ствола, которое придает металлу дополнительную прочность. Снаружи ружье покрыто никелем (хорош против ржавчины). Неудивительно, что производитель дает гарантию на то, что ИЖ-27 совершит не меньше 7000 выстрелов (по 3500 на ствол), прежде чем у него появятся какие-то проблемы.
к содержанию ↑
Модификации ИЖ-27 (МР-27)
Baikal MP-27 — именно под таким обозначением стал выпускаться ИЖ-27 после 2009 года. Помимо изменения в названии, в новой версии было также улучшено качество мелких деталей — теперь все они использовали сталь А50 и отличались длительным сроком службы. Однако большая часть модельного ряда — это образцы, вышедшие еще в Советском Союзе. Вот лишь самые популярные из ружей:
Однако большая часть модельного ряда — это образцы, вышедшие еще в Советском Союзе. Вот лишь самые популярные из ружей:
- ИЖ-27М (MP-27М) — одна из первых модификаций, увидевшая свет еще в 80-х годах прошлого столетия. Ее главное отличие от предшественника — наличие стандартной системы извлечения гильзы (вместо эжекторной), а также сразу два спусковых крючка для стрельбы (чок и получок).
- ИЖ-27Е (MP-27E) — видоизмененная версия стандартного ружья, главное отличие которого состоит в наличии более качественного эжектора. Многие покупатели жаловались, что иногда выбрасыватель гильз отказывался работать — это было исправлено в новой модели.
- ИЖ-27ЕМ (MP-27EM) — эта модель включает в себя все лучшее, что получило два предыдущих образца. У новой версии появилась возможность отключать эжектор с помощью специальной кнопки в случае необходимости.
- ИЖ-27М-1С (MP-27M-1C) — многим любителям охотничьего промысла не понравился тот факт, что современные модификации выходят в варианте с двумя курками.
 Для таких людей было принято решение разработать эту модель с переключателем боя в виде флажка.
Для таких людей было принято решение разработать эту модель с переключателем боя в виде флажка.
- ИЖ-27ЕМ-1С (MP-27EM-1C) — модифицированная версия ИЖ-27ЕМ с одним спусковым крючком. Как и в предыдущем случае переключение осуществляется с помощью специального механизма, расположенного на ствольной коробке.
- ИЖ-27М (MP-27M «Junior») — облегченная версия классической модификации, подходящая для подростков. Максимальный калибр выпуска ружья — 16-й. Подойдет для тренировок и спорта.
- ИЖ-27СТ — особая модификация «Юниора», заточенная под стендовую стрельбу. Основное отличие от оригинала — наличие усиленного чока, а также более длинного ствола — 760 мм. Выпуск этой версии прекращен ввиду ее ненадобности.
- ИЖ-27СК — спортивная модификация «Юниора», заточенная под круглый стенд. Длина ствола этой версии составляет всего 660 миллиметров, а калибр не может быть больше 20-ого. Выпуск модели сегодня прекращен.
- ИЖ-27 «Спортинг» — весьма распространенная модификация среди любителей соревновательной и тренировочной стрельбы.
 Ее главное отличие от других спортивных версий заключается в неограниченном выборе калибра, а также том, что ружье до сих пор выпускается ИЖМЕХОМ.
Ее главное отличие от других спортивных версий заключается в неограниченном выборе калибра, а также том, что ружье до сих пор выпускается ИЖМЕХОМ.
И это лишь те модификации, которые продаются чаще остальных. Не стоит забывать об уже упомянутых штучных ружьях или особых сувенирных образцах, к примеру, ИЖ-27 «РУСЬ», которое в 3 раза дороже серийного варианта. Отличить эксклюзивные модели можно по внешнему виду и увеличенному гарантийному сроку. Также УСМ в штучных и коллекционных модификациях отличается большей надежностью.
к содержанию ↑
Список штучных моделей
Эксклюзивные образцы ИЖ-27 в основном популярны среди коллекционеров и ценителей прекрасного. Учитывая то, что позволить себе такое ружье может отнюдь не каждый житель России, за все время существования ИЖМЕХ выпустил не так много подобных образцов. Вот краткое описание трех самых популярных версий:
- ИЖ-27-Ш1 — первая штучная модификация ружья ИЖ-27.
 Распознать ее можно по серебряной гравировке, находящейся на ствольной коробке ружья, а также более темному прикладу, изготовленному из красного дерева.
Распознать ее можно по серебряной гравировке, находящейся на ствольной коробке ружья, а также более темному прикладу, изготовленному из красного дерева. - ИЖ-27-Ш2 — вторая версия штучной модификации, которая отличается от первой золотым курком, а также наличием позолоченных вставок. Более дорогой, но куда более красивый вариант, поставляющийся в особом кейсе.
- ИЖ-27 «РУСЬ» — версия ИЖ-27, которой хотел бы обладать каждый коллекционер. От стандартного оружия она отличается гравировками и черным вороненым стволом. Одним из обладателей «Руси» является В. В. Путин.
Приобрести штучные версии ИЖ-27 сегодня можно только с рук у владельцев. ИЖМЕХ прекратил выпуск дорогого оружия ввиду его небольшой популярности, а исключения делает только по особым случаям. Из-за этой особенности сложно представить, во сколько покупателю обойдется такое оружие.
Из-за этой особенности сложно представить, во сколько покупателю обойдется такое оружие.
к содержанию ↑
Независимо от того, насколько оружие легендарно, нельзя забывать о его главных достоинствах и недостатках. Эта информация позволит избежать неприятных ситуаций после покупки или же поможет подобрать модификацию, которая подходит под предпочтения охотника. Начать перечисление лучше всего с преимуществ, поскольку их гораздо больше:
- Универсальность. Ружье ИЖ-27 не имеет узкой направленности, поэтому его могут использовать не только охотники, но и спортсмены и даже подростки. В зависимости от того, какой калибр оружия будет выбран, технические характеристики модели будут отличаться. Ружье ИЖ-27 может использовать патроны с различной навеской пороха и дроби, так что оно одинаково эффективно подходит как для охоты на пернатую дичь, так и для отстрела хищников.
- Эргономика. Основная проблема большинства ружей 12 калибра заключается в высокой отдаче, которая зачастую причиняет сильный дискомфорт и неудобства охотнику.
 Однако поскольку в модели ИЖ-27 используется более удобный приклад и ложе, а также имеется резиновый затыльник, инерция от выстрела является не такой сильной. Это позволяет увеличивать навеску дроби и использовать более мощные пороха.
Однако поскольку в модели ИЖ-27 используется более удобный приклад и ложе, а также имеется резиновый затыльник, инерция от выстрела является не такой сильной. Это позволяет увеличивать навеску дроби и использовать более мощные пороха.
- Безопасность. Оружие ИЖ-27 обладает трехступенчатой системой защиты — запирателем ствола, автоматическим предохранителем и механическим предохранителем. Первый уровень не дает сделать выстрел, если оружие закрыто не полностью. Второй — служит для защиты охотника и окружающих от случайного нажатия на спусковой крючок. Третий — устранит вероятность несчастного случая, связанную с падением ружья.
- Внешний вид. Штучные ружья ИЖ-27 выглядят действительно дорого и красиво.
 Использовать такое оружие на охоте — не слишком удачная идея, но в качестве коллекционной модели оно подходит идеально. Да и обычный ИЖ-27 также выглядит отлично — деревянные и металлические части соединены между собой крепко-накрепко, а чернение ствола выполнено ровно и однотонно.
Использовать такое оружие на охоте — не слишком удачная идея, но в качестве коллекционной модели оно подходит идеально. Да и обычный ИЖ-27 также выглядит отлично — деревянные и металлические части соединены между собой крепко-накрепко, а чернение ствола выполнено ровно и однотонно.
Из недостатков ружья большинство владельцев чаще всего выделяют большую массу модели (от 3,7 до 4 кг). Использовать такое громоздкое оружие для тренировки или охоты бывает затруднительно — человек попросту устает таскать с собой такую тяжесть или стрелять из нее. Хотя для взрослого мужчины средней комплекции проблема не является слишком большой.
к содержанию ↑
Как выбрать: новое или БУ
Эксклюзивная модель ИЖ-27 в кейсеОхотники также нередко задумываются, какое ружье ИЖ-27 им стоит купить: бывшее в употреблении или новое? Так или иначе для покупателя есть свои преимущества и недостатки. К примеру, б/у модель стоит меньше, однако есть риск стать обладателем оружия с проблемами. Оружейные магазины дают гарантию на товар, который реализуют, но делают это по более высокой цене. Так что вот вам несколько рекомендаций, позволяющих купить ИЖ-27 по низкой цене:
Оружейные магазины дают гарантию на товар, который реализуют, но делают это по более высокой цене. Так что вот вам несколько рекомендаций, позволяющих купить ИЖ-27 по низкой цене:
- Покупая оружие с рук, смотрите на его внешнее состояние. Ружье, которое использовалось редко, а также регулярно подвергалось чистке и смазке, никогда не будет иметь расшатанное ложе или приклад. Если же деревянные части шатаются — это признак того, что оружие все время работало на пределе, поэтому у него могут быть проблемы и с другими частями, например, УСМ.
- Приобретая оружие в магазине, не забывайте о своих правах покупателя. К примеру, любой товар можно вернуть в течение 2-х недель при предъявлении чека и коробки. Этим можно воспользоваться в том случае, если купленный ИЖ-27 не понравится по каким-то критериям. А уж если попалась бракованная модель, то вернуть ее в магазин и забрать свои деньги обратно будет легко.
- Независимо от того, покупаете ли вы ружье в магазине или с рук, любой добросовестный продавец не откажет покупателю в просьбе провести неполную разборку оружия.
 Это может быть отличной возможностью взглянуть на состояние всех механизмов модели. Особое внимание следует обратить на то, не растянуты ли пружины в УСМ, а также взглянуть на состояние патронника.
Это может быть отличной возможностью взглянуть на состояние всех механизмов модели. Особое внимание следует обратить на то, не растянуты ли пружины в УСМ, а также взглянуть на состояние патронника.
Рекомендуется отправляться за покупкой ружья вместе с другом, который хотя бы немного разбирается в оружии. Даже если вы оба не заметите какой-то недостаток, товарищ всегда сможет отговорить вас от сомнительной сделки. Если же пойти за покупкой в одиночку, то всегда будет соблазн купить ружье, даже если оно не в лучшем качестве, ведь это может привести к исполнению заветного желания.
к содержанию ↑
Хранение и уход
Очень часто владельцы совершают одни и те же распространенные ошибки, касающиеся его правильного хранения и ухода, что приводит к уменьшению срока эксплуатации, появлению различных поломок, а также снижению тактико-технических характеристик. Вот рекомендации, которые позволят избежать такого сценария:
- чистить оружие после каждого его использования — и не важно, сколько выстрелов вы сделали (10 или 100) — на стволе все равно будет образовываться освинцованный налет;
- хранить ружье только в сухом помещении, разместив его дулом вверх — это позволит избежать вытекания масла;
- чистить ствол стальным ершиком только в случае крайней необходимости — этот материал царапает хромированное покрытие;
- использовать керосин для устранения остатков старого масла — достаточно смочить в нем кусочек ветоши и насадить ее на шомпол;
- вымывать и высушивать ствол перед тем, как нанести на него новое масло — в противном случае качество смазки заметно ухудшится;
- не перебарщивать с маслянистым составом — если из ствола будет идти слишком много дыма, это может привести к ухудшению его состояния.

Каждый владелец ружья должен помнить, что чем лучше он ухаживает за своим оружием, тем дольше оно прослужит. И, несмотря на то, что ИЖ-27 считается неприхотливой моделью в плане обслуживания, пренебрегать основными правилами по уходу не следует.
к содержанию ↑
Тюнинг
Оружие ИЖ-27 хорошо еще и потому, что поддается модификациям в домашних условиях. Вот список популярных апгрейдов, которые проводят владельцы ружья чаще всего:
- Установка коллиматорного прицела вместо стандартной мушки и целика на планку Вивера.
- Замена деревянного ложа и приклада на пластиковые аналоги.
- Укорочение ствола для повышения мощности (страдает кучность боя).
- Использование различных дульных насадок для повышения кучности стрельбы.

- Крепление подствольного «обвеса» (штык-нож, фонарик, лазерный целеуказатель).
- Нанесение именной гравировки на дерево или металл.
- Окрашивание приклада и ложе или полировка лаком.
- Дополнительное чернение металлических деталей.
И это лишь самые популярные способы модификации ИЖ-27. При желании и наличии достаточного количества средств ружье можно изменить до неузнаваемости.
к содержанию ↑
Отзывы охотников
Большинство отрицательных отзывов вокруг ИЖ-27 базируются на том, что ружье громоздкое. Охотники жалуются, что ходить по лесу с 4-килограммовым агрегатом за спиной слишком утомительно. То ли дело импортные образцы, которые пусть и уступают российскому оружию по качеству, но зато весят меньше. Хотя с этим мнением согласны не все.
Хотя с этим мнением согласны не все.
Также некоторые охотники отмечают, что модель не слишком хорошо подходит для стрельбы патронами типа «магнум» с увеличенной навеской «Сокола». После использования такого боеприпаса металлические детали оружия начинали постепенно расшатываться, из-за чего приходилось задумываться об использовании дополнительных муфт и штифтов.
В целом владельцы ИЖ-27 крайне положительно отзываются о своем ружье. Форумчане отмечают, что даже б/у модель соответствует заявленному производителем качеству и характеристикам. Самое главное — не увлекаться экспериментами с различными навесками пороха и дроби в патроне, иначе даже малейшая ошибка в расчетах может привести к печальному исходу.
ИЖ-27 — это одно из самых популярных ружей от Ижевского завода, которое известно не только на территории нашей страны, но и за рубежом. Модель продолжает покупаться даже спустя почти 50 лет после выхода за счет качественной сборки, отличных ТТХ и приемлемой цены. А учитывая то, что ИЖМЕХ до сих пор поддерживает ружье выпуском различных модификаций, перспектива популярности ИЖ-27 и через 100 лет выглядит все более оптимистичной.
А учитывая то, что ИЖМЕХ до сих пор поддерживает ружье выпуском различных модификаций, перспектива популярности ИЖ-27 и через 100 лет выглядит все более оптимистичной.
БАЙКАЛ ИЖ 27 Новые и подержанные Цена, стоимость и тенденции 2022
- Главная
- БАЙКАЛ ИЖ 27
Поиск
Выберите категориюПистолетВинтовкаДробовикОптикаЖурналКатегория
- Сводка
- На продажу [1]
- Продано [33]
- Данные
Сколько стоит ружье БАЙКАЛ ИЖ 27?
Ружье БАЙКАЛ ИЖ 27 в настоящее время стоит средняя цена 510,00 долларов США. . Средняя цена за 12 месяцев составляет 510 долларов США.
Подержанная стоимость ружья БАЙКАЛ ИЖ 27 за последнее время выросла на 31,71 доллара. 12 месяцев по цене 510,00 долларов США.
Спрос на новые ружья БАЙКАЛ ИЖ 27 за последние 12 месяцев не изменился.
Спрос на подержанные ружья БАЙКАЛ ИЖ 27 за последнее время упал на 1 единицу. 12 месяцев.
12 месяцев.
Расчетная стоимость
*Использование условия 80% для расчета используемых значений.
* Калибр, длина ствола, поколения, подмодели могут влиять на цену предмета.
Убедитесь, что ваш поиск достаточно конкретен, чтобы получить правильное значение.
| Используется | Новый | |
|---|---|---|
| Торговля | 331,50 $ | — |
| Частная вечеринка | 510,00 $ | — |
Хотите продать свое оружие?
Продай ружье БАЙКАЛ ИЖ 27
Получить предложение
Ружье БАЙКАЛ ИЖ 27 Продам
БАЙКАЛ ИЖ-27ЭМ GUNS.COM
$650.99 Просмотреть сайт
Просмотреть все Продажа
Недавно продано Ружье БАЙКАЛ ИЖ 27
| Цена | Товар | Состояние | Дата продажи |
|---|---|---|---|
| 320,00 $ | BAIKAL KBI МОДЕЛЬ ИЖ-27М 12 КАЛИБРОВ ПОД РУЖЬЕМ $1 БЕЗ РЕЗЕРВА 28 ДЮЙМОВЫЙ СТВОЛ Andover, CT 06232 | Используется | |
| 595,00 $ | БАЙКАЛ 12 КАМЕРА ИЖ-27ЭМ-1С 28 ДЮЙМОВЫЕ СТВОЛЫ ДЕЙСТВИТЕЛЬНО ХОРОШЕЕ ДЕРЕВО! Вест-Пойнт, NE 68788 | Используется | |
| 300,00 $ | 12 КАЛИБРОВКА БАЙКАЛ (EAA) МОДЕЛЬ ИЖ-27ЕМ-1С O/U 12GA РУЖЬЕ Лайонс, Иллинойс | Используется | |
| 600,00 $ | IZH-27 BAIKAL IZH 27EM-1C IZH37EM1C 12 GA 3″ ДУЛЬНАЯ НАСАДКА 26,25″ НАД/НИЖНЯЯ РУЖЬЯ Simi Valley, CA | Используется | |
| 400,00 $ | 12 КАЛИБРОВКА БАЙКАЛ (EAA) МОДЕЛЬ ИЖ-27ЕМ-1C O/U 12GA РУЖЬЕ Lyons, IL | Используется | |
| 417,00 $ | РУЖЬЕ ИЖ-27 БАЙКАЛ МОДЕЛЬ ИЖ37 12 КАЛИБРА ВЕРТИКАЛЬНОЕ РУЖЬЕ Martinsburg, WV 25405 | Используется | |
| 499,95 $ | РУЖЬЕ ИЖ-27 БАЙКАЛ МОДЕЛЬ ИЖ37 12 КАЛИБРОВЫЙ ВЕРТИКАЛЬНЫЙ ДРОБОВИК, 28 ДЮЙМОВЫЙ СТВОЛ Martinsburg, WV 25405 | Используется | |
| 465,00 $ | BAIKAL МОДЕЛЬ IZH-27EM-1C-M O/U 12 КАЛИБРОВКА В ОТЛИЧНОМ СОСТОЯНИИ 28 ДЮЙМОВЫЙ СТВОЛ Wales Center, NY 14169 | Используется |
- Б/у Продано [33]
- Новый Продан [0]
НОВЫЙ БАЙКАЛ ИЖ 27 ЦЕНА
Хотите продать свое ружье?
Продай ружье БАЙКАЛ ИЖ 27
Получить предложение
Нет результатов.
Б/У БАЙКАЛ ИЖ 27 ЦЕНА
Хотите продать свое ружье?
Продай ружье БАЙКАЛ ИЖ 27
Получить предложение
Байкал КБИ Модель ИЖ-27М 12 Калибр Верхний Под Ружье $1 Без Резерва Ствол 28 дюймов
ЦЕНА:
$320. 00 00 | ПРОИЗВОДИТЕЛЬ: Байкал |
| СОСТОЯНИЕ: Используется | МОДЕЛЬ: ИЖ-27 |
| ПРОДАНО: | СКП: |
| РАСПОЛОЖЕНИЕ: Андовер, Коннектикут, 06232. | Артикул: |
| КАЛИБР: | РУЧН. ЧАСТЬ #: |
| ВМЕСТИМОСТЬ: 2 | ДЛИНА СТВОЛА: 28 дюймов |
Байкал 12 калибра ИЖ-27ЕМ-1С 28-дюймовые стволы ДЕЙСТВИТЕЛЬНО ХОРОШЕЕ ДЕРЕВО!
| ЦЕНА: $595,00 | ПРОИЗВОДИТЕЛЬ: Байкал |
| СОСТОЯНИЕ: Используется | МОДЕЛЬ: ИЖ-27 |
| ПРОДАНО: | СКП: |
| РАСПОЛОЖЕНИЕ: Вест-Пойнт, NE 68788. | Артикул: |
| КАЛИБР: | РУЧН. ЧАСТЬ #: |
| ВМЕСТИМОСТЬ: 2 | ДЛИНА СТВОЛА: 28 дюймов |
Байкал 12 калибра (EAA) Модель ИЖ-27ЕМ-1С О/У Ружье 12 калибра
ЦЕНА:
$300. 00 00 | ПРОИЗВОДИТЕЛЬ: Байкал |
| СОСТОЯНИЕ: Используется | МОДЕЛЬ: |
| ПРОДАНО: | СКП: |
| РАСПОЛОЖЕНИЕ: Лион, Иллинойс | Артикул: |
| КАЛИБР: 12 Калибр | РУЧН. ЧАСТЬ #: |
| ВМЕСТИМОСТЬ: | ДЛИНА СТВОЛА: |
ИЖ-27 Байкал ИЖ 27ЕМ-1С ИЖ37ЕМ1С 12 Ga 3″ Дроссельная трубка 26,25″ Верхнее/нижнее ружье
| ЦЕНА: 600,00 долларов США | ПРОИЗВОДИТЕЛЬ: Байкал |
| СОСТОЯНИЕ: Используется | МОДЕЛЬ: ИЖ-27 |
| ПРОДАНО: | СКП: |
| РАСПОЛОЖЕНИЕ: Сими-Вэлли, Калифорния | Артикул: |
| КАЛИБР: | РУЧН. ЧАСТЬ #: |
| ВМЕСТИМОСТЬ: | ДЛИНА СТВОЛА: |
Байкал 12 калибра (EAA) Модель ИЖ-27ЕМ-1С O/U Ружье 12ga
| ЦЕНА: 400,00 долларов США | ПРОИЗВОДИТЕЛЬ: Байкал |
| СОСТОЯНИЕ: Используется | МОДЕЛЬ: |
| ПРОДАНО: | СКП: |
| РАСПОЛОЖЕНИЕ: Лион, Иллинойс | Артикул: |
| КАЛИБР: 12 Калибр | РУЧН. ЧАСТЬ #: |
| ВМЕСТИМОСТЬ: | ДЛИНА СТВОЛА: |
Ружье ИЖ-27 Байкал Модель ИЖ37 Калибр 12 Вертикальное ружье
| ЦЕНА: 417,00 долларов США | ПРОИЗВОДИТЕЛЬ: Байкал |
| СОСТОЯНИЕ: Используется | МОДЕЛЬ: ИЖ-27 |
| ПРОДАНО: | СКП: |
| РАСПОЛОЖЕНИЕ: Мартинсбург, WV 25405 | Артикул: |
| КАЛИБР: | РУЧН. ЧАСТЬ #: |
| ВМЕСТИМОСТЬ: | ДЛИНА СТВОЛА: |
ИЖ-27 Байкал Модель ИЖ37 Калибр 12 Вертикальное ружье Ствол 28 дюймов
| ЦЕНА: 499,95 долларов США | ПРОИЗВОДИТЕЛЬ: Байкал |
| СОСТОЯНИЕ: Используется | МОДЕЛЬ: ИЖ-27 |
| ПРОДАНО: | СКП: |
| РАСПОЛОЖЕНИЕ: Мартинсбург, WV 25405 | Артикул: |
| КАЛИБР: | РУЧН. ЧАСТЬ #: |
| ВМЕСТИМОСТЬ: | ДЛИНА СТВОЛА: 28 дюймов |
БАЙКАЛ МОДЕЛЬ ИЖ-27ЕМ-1С-М O/U 12 Калибр Отличное Состояние 28 дюймов «ствол
| ЦЕНА: 465,00 долларов США | ПРОИЗВОДИТЕЛЬ: Байкал |
| СОСТОЯНИЕ: Используется | МОДЕЛЬ: ИЖ-27 |
| ПРОДАНО: | СКП: |
| РАСПОЛОЖЕНИЕ: Уэльс-центр, NY 14169 | Артикул: |
| КАЛИБР: | РУЧН. ЧАСТЬ #: |
| ВМЕСТИМОСТЬ: 2 | ДЛИНА СТВОЛА: 28 дюймов |
Байкал Россия Модель ИЖ-27ЕМ-1C Ствол 20 GA 26 дюймов 26 дюймов
| ЦЕНА: $500.00 | ПРОИЗВОДИТЕЛЬ: Байкал |
| СОСТОЯНИЕ: Используется | МОДЕЛЬ: ИЖ-27 |
| ПРОДАНО: | СКП: |
| РАСПОЛОЖЕНИЕ: Конроу, Техас, 77384. | Артикул: |
| КАЛИБР: | РУЧН. ЧАСТЬ #: |
| ВМЕСТИМОСТЬ: 2 | ДЛИНА СТВОЛА: 26 дюймов |
ИЖ-27 Спортинг Русский Байкал ИЖ-27ЕМ-1С 12 Калибр больше/меньше 3 дюйма с отверстиями 29-1/2 дюйма Стволы
| ЦЕНА: 600,00 долларов США | ПРОИЗВОДИТЕЛЬ: Байкал |
| СОСТОЯНИЕ: Используется | МОДЕЛЬ: ИЖ-27 Спортинг |
| ПРОДАНО: | СКП: |
| РАСПОЛОЖЕНИЕ: Франклин, Пенсильвания, 16323. | Артикул: |
| КАЛИБР: | РУЧН. ЧАСТЬ #: |
| ВМЕСТИМОСТЬ: | ДЛИНА СТВОЛА: |
Ружье ИЖ-27 Байкал ИЖ 27ЕМ EAA Corp., ствол 28 дюймов
| ЦЕНА: $581,98 | ПРОИЗВОДИТЕЛЬ: Байкал |
| СОСТОЯНИЕ: Используется | МОДЕЛЬ: ИЖ-27 |
| ПРОДАНО: | СКП: |
| РАСПОЛОЖЕНИЕ: Гудзон, Висконсин, 54016 | Артикул: |
| КАЛИБР: | РУЧН. ЧАСТЬ #: |
| ВМЕСТИМОСТЬ: 2 | ДЛИНА СТВОЛА: 28 дюймов |
Русский Байкал ИЖ-27ЕМ-1С 12 Калибр больше/меньше 28-1/2” вороненый с коробкой
| ЦЕНА: $530.00 | ПРОИЗВОДИТЕЛЬ: Байкал |
| СОСТОЯНИЕ: Используется | МОДЕЛЬ: ИЖ-27 |
| ПРОДАНО: | СКП: |
| РАСПОЛОЖЕНИЕ: Франклин, Пенсильвания, 16323. | Артикул: |
| КАЛИБР: | РУЧН. ЧАСТЬ #: |
| ВМЕСТИМОСТЬ: | ДЛИНА СТВОЛА: |
Русский Байкал ИЖ-27ЕМ-1С 12 калибра больше/меньше 29-1/2” с портовым стволом
| ЦЕНА: $525,00 | ПРОИЗВОДИТЕЛЬ: Байкал |
| СОСТОЯНИЕ: Используется | МОДЕЛЬ: ИЖ-27 |
| ПРОДАНО: | СКП: |
| РАСПОЛОЖЕНИЕ: Франклин, Пенсильвания, 16323. | Артикул: |
| КАЛИБР: | РУЧН. ЧАСТЬ #: |
| ВМЕСТИМОСТЬ: | ДЛИНА СТВОЛА: |
Русский Baikal IZH-27EM-1C Больше/меньше 12 калибра 28-1/2” с коробкой 3 чока
| ЦЕНА: $500.00 | ПРОИЗВОДИТЕЛЬ: Байкал |
| СОСТОЯНИЕ: Используется | МОДЕЛЬ: ИЖ-27 |
| ПРОДАНО: | СКП: |
| РАСПОЛОЖЕНИЕ: Франклин, Пенсильвания, 16323. | Артикул: |
| КАЛИБР: | РУЧН. ЧАСТЬ #: |
| ВМЕСТИМОСТЬ: | ДЛИНА СТВОЛА: |
Русский Байкал ИЖ-27ЕМ-1С Больше/Меньше 12 калибра с коробкой 29Стволы -1/2”
| ЦЕНА: $530.00 | ПРОИЗВОДИТЕЛЬ: Байкал |
| СОСТОЯНИЕ: Используется | МОДЕЛЬ: ИЖ-27 |
| ПРОДАНО: | СКП: |
| РАСПОЛОЖЕНИЕ: Франклин, Пенсильвания, 16323. | Артикул: |
| КАЛИБР: | РУЧН. ЧАСТЬ #: |
| ВМЕСТИМОСТЬ: | ДЛИНА СТВОЛА: |
Хотите продать свое оружие?
Продай ружье БАЙКАЛ ИЖ 27
Получить предложение
Средняя новая цена
—
Средняя подержанная цена 0
Всего новых проданных
—
Всего использовано Продано 0
Манометр/Калибр. .410 Диаметр ствола 12 млрд лет 16 млрд лет 20 года 28 года Все варианты… Конфигурация ствола Одинокий Бок о бок Больше-меньше Комбо Все варианты… Курок Одинокий Двойной Золото Все варианты… Год Все атрибуты… Байкал Ремингтон Браунинг Армс S&W Винчестер Все производители… Дробовик
Винтовка
Пистолет
Все типы. Мобильные телефоны и мобильные устройства Компьютеры Музыкальные инструменты Аудио оборудование Оружие Видео и ТВ Винтаж Профессиональный звук Оборудование для камеры Копировальные аппараты Электроинструмент Видеоигры Автомобильная стереосистема |
Результаты поиска 1–29 < предыдущая - Страница: 1 - следующий >
< предыдущая - Страница: 1 - следующий > | Гитарная компания Fowlcon Стаутон доц. Хобби Семейный залог и ссуда КалибрыСША Шумарт Лимитед Бил Ассошиэйтс действующий военный инвалид Брикер ТК Услуги эпический Фармацевт на пенсии Распрехер Название компании Пограничный обмен | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 ..
.. 410 (36GA), 12 Ga, 16 Ga, 20 Ga — Длина ствола: 26 дюймов, 28 дюймов — Конфигурация ствола: одинарный — Дроссель: индейка
410 (36GA), 12 Ga, 16 Ga, 20 Ga — Длина ствола: 26 дюймов, 28 дюймов — Конфигурация ствола: одинарный — Дроссель: индейка 410 (36GA), 12 Ga, 20 Ga — Длина ствола: 26 дюймов, 28 дюймов — Конфигурация ствола: одинарная
410 (36GA), 12 Ga, 20 Ga — Длина ствола: 26 дюймов, 28 дюймов — Конфигурация ствола: одинарная 410 (36GA), 20 Ga — Затвор: BoxLock — Материал рукоятки/ложи: Орех — Стиль рукоятки/ложи: Клетчатый — Длина ствола: 26,5 дюйма — Конфигурация ствола: Side-by-Side — Материал ствола: Сталь — Отделка ствола: воронение — Спусковой крючок: одиночный — Дроссель: полный, модифицированный
410 (36GA), 20 Ga — Затвор: BoxLock — Материал рукоятки/ложи: Орех — Стиль рукоятки/ложи: Клетчатый — Длина ствола: 26,5 дюйма — Конфигурация ствола: Side-by-Side — Материал ствола: Сталь — Отделка ствола: воронение — Спусковой крючок: одиночный — Дроссель: полный, модифицированный 410 (36GA), 12 Ga, 20 Ga, 28 Ga — Конфигурация ствола: Side-by-Side
410 (36GA), 12 Ga, 20 Ga, 28 Ga — Конфигурация ствола: Side-by-Side
 Инк
Инк Ружье Байкал ИЖ-27ЕМ-1С O/U калибра 20 — 22.
 08.2020
08.2020Ружье Байкал ИЖ-27ЕМ-1C O/U 20 калибра
Аналогичная история продаж
Просмотреть больше товаров в Оружие и огнестрельное оружиеПродано
2022НОВИНКА В КОРОБКЕ JTS M12AK ПОЛУАВТОМАТИЧЕСКИЙ РУЖЬЕ 12 КАЛ.См. Цена продажи
Продано
2022MOSSBERG МОДЕЛЬ 835 ПОМПОВОЕ РУЖЬЕ 12 КАЛ.См. Цена продажи
Проданный
202222222.radikal ARM NK-1 BULLPUP 24IN 12 калибра.
20221991 MOSSBERG MODEL 590 12 GA ПОМПОВОЕ РУЖЬЕСмотреть цену продажи
Продано
2022Remington 870 Express Super Magnum Slug Gun Помповое ружьеСмотреть цену продажи
Sold
2022AYA/Firearms International SxS ShotgunSee Sold Price
Sold
2022AMERICAN TACTICAL BULLDOG SEMI AUTO 12GA SHOTGUNSee Sold Price
Sold
2022Smith & Wesson Model 1000 Semi Automatic ShotgunSee Sold Price
Sold
2022Savage Fox Sterlingworth
2022Neumann Bros. Percussion ShotgunСм. Цена
Percussion ShotgunСм. Цена
2022ROCK ISLAND ARMORY MERIVA 12GA ПОМПОВОЕ РУЖЬЕСм. Цена
Проданный
202222 Остров Остров Остров Армьера Мерива 12GA Действие Действие Действие Действие Действие Проданная цена
Проданная
2022 Амантино/Stoeger Single Shotgunsee Sald Prode
Проданный
2022nepalese Gahendra Martina Martini Rifles. Ранние английские двуствольные ружья
350 $1 октября 2022 г. (M) COCA-COLA CENTENNIAL WINCHESTER MODEL 1500 XTR ПОЛУАВТОМАТИЧЕСКОЕ РУЖЬЕ С КОРОБКОЙ.
$950Осталось 2 дня19ДВУСТОРОННЕЕ УДАРНОЕ РУЖЬЕ TH CENT
160 $ 1 день Осталось (2) универсал старые винтовки, вкл. Двуствольное ружье U S Springfield 1822 & Richards
Двуствольное ружье U S Springfield 1822 & Richards
Доступно для покупки Модели Fine Art Масштаб 1:32 (калибр 1) ЛАТУНЬ Pennsylvania M1a 4-8-2 Steam Loco & Tender, c10 в коробке c10,
7 996 $03 октября 2022 г. AMERICAN TACTICAL Дробовик CAVALRY 20 GAUGE NIB
230 $03 октября 2022 г. MOSSBERG 500A 12 GAUGE SHOTGUN & THREE BARRELS
$ 13003 октября 2022 г.WINCHESTER MODEL 12 20 GAUGE SHOTGUN
90 долларов США29 сентября 2022 г. WINCHESTER MODEL 1200 12 КАЛИБРОВОЕ РУЖЬЕ.
$ 200Октябрь 03, 2022J Стивенс Браунинг Действие Насоса 12 GA
$ 50ОКТ 03, 2022 Кхан Catamount Arms Lynxx 12 калибровка ружья
$ 50Oct 23, 2022 Mossberg 500. .410 Pump Shotgun
$500 3 октября 2022 г. MOSSBERG 500A 12 GAUGE PUMP ACTION SHOTGUN
MOSSBERG 500A 12 GAUGE PUMP ACTION SHOTGUN
03 октября 2022 г.80 $
23 октября 2022 г. Mossberg 590 .410 Gauge Pump Action Shotgun
125 $Другие товары в Оружие и огнестрельное оружие
Просмотреть еще1 день Осталось (2) Estate old guns incl. США Двуствольное ружье Springfield 1822 & Richards
400 долларов США01 октября 2022 г. (M) COCA-COLA CENTENNIAL WINCHESTER MODEL 1500 XTR ПОЛУАВТОМАТИЧЕСКОЕ РУЖЬЕ С КОРОБКОЙ.
950 $Осталось 2 дня УДАРНОЕ РУЖЬЕ 19 CENT
160 $ Осталось 7 дней Три раннеанглийских двуствольных ружья
350 $30 сентября 2022 г. Ca 1810 Британский мушкет с башенным замком
100 долл. США 01 окт. 2022 г. Отличная пара корабельных пушек Блумфилда Королевского флота Англии, около 1780 г.
5 дней Вешалка
$ 55Октябрь 02, 2022SPRINGFIELD 1837 Musket
$ 350Октябрь 02, 2022 Мантон из Калькутты -Боуи Нож
$ 250Октябрь 01, 2022 Lalgge 19 -го века US Bowie Knife с Edged Blade
$ 2009октябрь 22, 2022C. 1890 Ножны и ремень с пряжками Crow Beaded
$50022 октября 2022 г.C. 1880 Nez Perce, расшитый бисером, в ножнах и торговом ноже
$800Осталось 3 часаБольшая латунная деревянная коробка Continental
$150022 октября 2022 г. Western Cowboy Кожаные седельные сумки c. 1960-е
125 $2 дня Left19th C. USA Военные латунные пуговицы, пули и колпачки (15)
275 $Рекомендуемое оружие и доспехи
Просмотреть еще2 days LeftWWII Немецкий люфтваффе с гравировкой Weyersberg Dagger
525 $ 1 день Left(1 day Left) старые винтовки в т. ч. US Springfield 1822 и двуствольное ружье Richards
ч. US Springfield 1822 и двуствольное ружье Richards
3 дня левого джапонского Samurai Sword
$ 10002 дня слева 20th C. Argentine Gaucho Facon Ножи 240
$ 7 дней LeftTintype of Civil War Confederate Soldier
5506 дней LeftA Lega Initiation Knife
CHF 1602 days LeftWWII Кинжал лидеров немецкого Красного Креста
$3903 days LeftWWII Парад японских офицеров с мечами и ножнами
100 $01 октября 2022 г. (M) COCA-COLA CENTENNIAL WINCHESTER MODEL 1500 XTR ПОЛУАВТОМАТИЧЕСКОЕ РУЖЬЕ С КОРОБКОЙ.
$ 9502 дня левтов Гражданская война Saber
$ 4002 дня левая 15 -й цент перкуссия двойной бочки
$ 1607 дней левой группы из 17 серебряных медалей
$ 3007 дней Leftthree Ранний английский Double Barrels
$ 350 Selecp 30, 20222 2CA Ранний английский. Мушкет с башенным замком
Мушкет с башенным замком
Подробная информация о товаре
Описание
Вертикальное ружье ИЖ-27ЭМ-1С производства Байкал в России под патрон 20 калибра с патронником 3 дюйма. Ружье имеет маркировку на нижней стороне ствольной коробки с обозначением модели. , название компании и импортер. Ружье имеет вороненую отделку ствола и ствольной коробки в хорошем состоянии с небольшим износом. Ложа и цевье в очень хорошем состоянии с красивыми насечками на цевье и запястье. Ствол имеет длину 28 дюймов и грязный канал ствола. Смотрите фото. СН 012704226 Б
Премия покупателя
- 20%
Эстимейт $300 — $500
22 августа 2020 г.
Стартовая цена
$150
0 регистрация.
Правила доставки, оплаты и аукциона
ИЖ 27 Руководство — [Документ в формате PDF]
МОДЕЛЬ EAA БАЙКАЛ ИЖ37
РУЖЬЕ ВЕРХНЕЕ И НИЖНЕЕ (12,
16, 20, 28 GA и . 9100RNS BOre 8 и .4100RNS РУКОВОДСТВО ПО ЭКСПЛУАТАЦИИ
9100RNS BOre 8 и .4100RNS РУКОВОДСТВО ПО ЭКСПЛУАТАЦИИ
European American Armory European American Armory Corp. 402 Ричард Роуд 3855 Северное шоссе США 1 Рокледж, Флорида. 32926 Какао, Флорида 32955 (321) 639-4842 www.eaacorp.com Электронная почта: [email protected] Copyright 2004
EAA CORP. МОДЕЛЬ: ИЖ37 ВЕРХНИЙ И НИЖНИЙ ПРЕДУПРЕЖДЕНИЯ И РУКОВОДСТВО ПО ЭКСПЛУАТАЦИИ ВНИМАТЕЛЬНО ПРОЧИТАЙТЕ ЭТУ ИНСТРУКЦИЮ НОМЕР МОДЕЛИ ИЖ37 ДЛИНА СТВОЛА 26″, 28″, 30″, 32″ ОТДЕЛКА СИНЯЯ КАЛИБРЫ 12, 20, 16, 28, 410
ТИП РУЖЬЯ ВЕРХ/НИЗ
ОБЩИЕ СВЕДЕНИЯ: Вес (средний) — 28 дюймов = 7,7 фунта. Длина — 43″, 45″, 47″, 49″
ДЛЯ ВАШЕЙ БЕЗОПАСНОСТИ И БЕЗОПАСНОСТИ ДРУГИХ, это руководство по эксплуатации
содержит важные предупреждения, инструкции и процедуры безопасности
которые необходимо понять ДО использования этого огнестрельного оружия. Читать ВСЕ
РУКОВОДСТВО, и если вы не понимаете какую-либо его часть или операцию
вашего ружья, остановитесь и обратитесь за профессиональной инструкцией по обращению с огнестрельным оружием. Вы можете получить копию этого руководства в European American Armory.
Корп. по запросу.
Вы можете получить копию этого руководства в European American Armory.
Корп. по запросу.
Запишите серийный номер вашего огнестрельного оружия (находится в верхней части ресивер под верхним рычагом): # _______________________________________________ и описание этого: ________________________________________________________
Сделайте копию данного руководства и подтверждение покупок/продаж запись на хранение в надежном сейфе или несгораемом контейнере. Такое доказательство необходимо, если огнестрельное оружие повреждено, украдено или отправлено в гарантийный ремонт.
2
ПРЕДУПРЕЖДЕНИЕ **ВЫ РИСКЕТЕ ПОЛУЧИТЬ ТРАВМУ ИЛИ СМЕРТЬ ПРИ РАБОТЕ С ЭТИМ
ОГНЕСТРЕЛЬНОЕ ОРУЖИЕ** Прежде чем обращаться с каким-либо огнестрельным оружием, убедитесь, что вы знаете
техники безопасности и, самое главное, держите огнестрельное оружие направленным в
безопасное направление. Прежде чем обращаться с этим огнестрельным оружием, прочтите, поймите
и следуйте инструкциям данного руководства. Всегда храните это руководство
с огнестрельным оружием. Если вы одалживаете, дарите или продаете это огнестрельное оружие, обязательно
руководство идет с ним. Если есть что-то, чего вы не делаете
понять, обратитесь за советом к специалисту по безопасному обращению с
огнестрельное оружие. Вы должны ВСЕГДА соблюдать следующие предупреждения, инструкции
и техники безопасности: 1. 2. Всегда держите дуло направленным в
безопасное направление. Огнестрельное оружие должно быть разряжено и закреплено, если оно не
фактически в использовании. Не полагайтесь на безопасность своего огнестрельного оружия. Будьте уверены в своем
цель и что за ней. Используйте только подходящие боеприпасы, соответствующие
точный калибр вашего конкретного ружья. Если ваш дробовик не
стрелять при нажатии на спусковой крючок, обращайтесь с ним осторожно. Всегда носить ухо
и защита глаз при стрельбе. Убедитесь, что в бочках нет
препятствия перед съемкой. Не переделывайте и не модифицируйте свое огнестрельное оружие и
обслуживайте его регулярно.
Всегда храните это руководство
с огнестрельным оружием. Если вы одалживаете, дарите или продаете это огнестрельное оружие, обязательно
руководство идет с ним. Если есть что-то, чего вы не делаете
понять, обратитесь за советом к специалисту по безопасному обращению с
огнестрельное оружие. Вы должны ВСЕГДА соблюдать следующие предупреждения, инструкции
и техники безопасности: 1. 2. Всегда держите дуло направленным в
безопасное направление. Огнестрельное оружие должно быть разряжено и закреплено, если оно не
фактически в использовании. Не полагайтесь на безопасность своего огнестрельного оружия. Будьте уверены в своем
цель и что за ней. Используйте только подходящие боеприпасы, соответствующие
точный калибр вашего конкретного ружья. Если ваш дробовик не
стрелять при нажатии на спусковой крючок, обращайтесь с ним осторожно. Всегда носить ухо
и защита глаз при стрельбе. Убедитесь, что в бочках нет
препятствия перед съемкой. Не переделывайте и не модифицируйте свое огнестрельное оружие и
обслуживайте его регулярно. Изучите механику и управление
характеристики вашего ружья. Стреляй трезвым!
Изучите механику и управление
характеристики вашего ружья. Стреляй трезвым!
3. 4. 5.
6.
7. 8.
9.
10.
11.
в этом огнестрельном оружии могут использоваться боеприпасы промышленного производства. Избегать военные излишки или старые боеприпасы, так как они могут не стрелять силы, достаточной для выталкивания снаряда или пыжа из ствола. Использование перезаряжаемых боеприпасов, боеприпасов высокого давления или ручного заряжания может быть опасно и приведет к аннулированию гарантии.
ПРЕДУПРЕЖДЕНИЕ О СВИНЦЕ: настоящее огнестрельное оружие не содержит свинца;
однако он стреляет боеприпасами, содержащими свинец или свинец.
соединения, известные в штате Калифорния как вызывающие рак, родовые
дефекты, репродуктивная токсичность и другие серьезные телесные повреждения.
Те, кто стреляет из огнестрельного оружия, встаньте рядом с тем, кто стреляет
огнестрельное оружие или чистое огнестрельное оружие настоящим предупреждаются об опасностях
представлены свинцом и соединениями свинца и должны принимать защитные
меры по охране здоровья. Избегайте воздействия свинца при обращении и мытье
Ваши руки после контакта. Надлежащая вентиляция абсолютно
необходимо при съемке в помещении.
Избегайте воздействия свинца при обращении и мытье
Ваши руки после контакта. Надлежащая вентиляция абсолютно
необходимо при съемке в помещении.
УВЕДОМЛЕНИЕ РУЖЬЯ КЛАССИФИЦИРУЮТСЯ КАК ОГНЕСТРЕЛЬНОЕ ИЛИ ОПАСНОЕ ОРУЖИЕ
и продаются European American Armory Corp.
понимая, что мы никоим образом не несем ответственности
для их безопасного обращения или перепродажи в соответствии с местными законами и правилами.
European American Armory Corp. не несет ответственности ни в
каким бы то ни было образом за неисправность огнестрельного оружия, за физическое
телесные повреждения или материальный ущерб, возникшие полностью или частично в результате
(1) преднамеренный или небрежный сброс, (2) неправильный или небрежный
обращении, (3) несанкционированных модификациях и/или изменении
внутренние/предохранительные механизмы, (4) дефектные, неподходящие, высокое давление,
снаряженные вручную или перезаряженные боеприпасы, (5) коррозия и/или отсутствие
надлежащее обслуживание (6) пренебрежение или (7) другие влияния, выходящие за рамки нашего
прямой и непосредственный контроль. Это ограничение действует независимо от
установлена ли ответственность на основании договора, небрежность
или строгая ответственность (включая любое отсутствие предупреждения). Под нет
обстоятельств, европейская американская оружейная корпорация будет нести ответственность за
случайные или косвенные убытки, такие как невозможность использования
имущества, коммерческих убытков и упущенной выгоды.
Это ограничение действует независимо от
установлена ли ответственность на основании договора, небрежность
или строгая ответственность (включая любое отсутствие предупреждения). Под нет
обстоятельств, европейская американская оружейная корпорация будет нести ответственность за
случайные или косвенные убытки, такие как невозможность использования
имущества, коммерческих убытков и упущенной выгоды.
4
СОБЛЮДЕНИЕ ЭТИХ ПРАВИЛ БЕЗОПАСНОСТИ МОЖЕТ СОХРАНИТЬ ВАШУ ЖИЗНЬ ИЛИ ЖИЗНЬ ДРУГИХ
КТО-ТО ДРУГОЙПройдите компетентный инструктаж по безопасному обращению с огнестрельным оружием
прежде чем использовать это огнестрельное оружие. Уточните у своего дилера названия
квалифицированные инструкторы. Все оружие чрезвычайно опасно, если
небрежно обращались или использовались. Помните, что пользователь огнестрельного оружия
безопасность и что полагаться на любое механическое устройство следует
никогда не заменяйте процедуры безопасного обращения. Это огнестрельное оружие БУДЕТ СТРЕЛЯТЬ, если
патрон в патроннике и спусковой крючок нажат, пока
безопасность отключена. НЕ ЗАРЯЖАЙТЕ ЭТО РУЖЬЕ, ПОКА НЕ ПОНИМАЕТЕ
КАК ЭТО РАБОТАЕТ. ДЕРЖИТЕ это ружье разряженным все время, кроме
когда вы готовы стрелять и уверены в своей цели. ВСЕГДА носить
это ружье в футляре или оригинальной коробке без патронов. НИКОГДА не указывайте на это
из дробовика во все, что не является вашей предполагаемой целью, даже если
ружье разряжено. При погрузке, разгрузке, очистке или обработке
всегда следите за тем, чтобы дуло было направлено в безопасном направлении, и всегда
держи палец на спусковом крючке. НИКОГДА не предполагайте, что какое-либо оружие разряжено.
Перед работой убедитесь, что камера пуста. Видеть
Инструкции по разгрузке. Убедитесь, что вы знаете, что это или любое огнестрельное оружие
выгружается перед отображением или обработкой. При обращении с любым
огнестрельного оружия, держите пальцы подальше от спускового крючка, пока
вы собираетесь стрелять. Получите инструкции от компетентного огнестрельного оружия
инструктору, прежде чем использовать это или любое огнестрельное оружие.
НЕ ЗАРЯЖАЙТЕ ЭТО РУЖЬЕ, ПОКА НЕ ПОНИМАЕТЕ
КАК ЭТО РАБОТАЕТ. ДЕРЖИТЕ это ружье разряженным все время, кроме
когда вы готовы стрелять и уверены в своей цели. ВСЕГДА носить
это ружье в футляре или оригинальной коробке без патронов. НИКОГДА не указывайте на это
из дробовика во все, что не является вашей предполагаемой целью, даже если
ружье разряжено. При погрузке, разгрузке, очистке или обработке
всегда следите за тем, чтобы дуло было направлено в безопасном направлении, и всегда
держи палец на спусковом крючке. НИКОГДА не предполагайте, что какое-либо оружие разряжено.
Перед работой убедитесь, что камера пуста. Видеть
Инструкции по разгрузке. Убедитесь, что вы знаете, что это или любое огнестрельное оружие
выгружается перед отображением или обработкой. При обращении с любым
огнестрельного оружия, держите пальцы подальше от спускового крючка, пока
вы собираетесь стрелять. Получите инструкции от компетентного огнестрельного оружия
инструктору, прежде чем использовать это или любое огнестрельное оружие. Учитесь у
профессионал, как обращаться, загружать, разгружать, эксплуатировать, стрелять и ухаживать
для вашего пистолета. НИКОГДА НЕ ИГРАЙТЕ со своим огнестрельным оружием. Это не игрушка и может
быть очень опасным оружием, которое может привести к серьезным травмам
или смерть. Всегда держите пистолет направленным в безопасном направлении. Удовольствие
каждое ружье как будто заряжено…все время. НИКОГДА не указывайте
огнестрельное оружие (заряженное или незаряженное) против любого человека или чего-либо, что вам не
намерен стрелять. Всегда держите ружье незаряженным, пока оно не будет готово к использованию.
и убедитесь, что он разряжен перед очисткой. Держи палец
ОТКЛЮЧАЙТЕ спусковой крючок до тех пор, пока вы не прицелитесь в цель и
готов стрелять. Научитесь удерживать палец вне спускового крючка
сторожить. Никогда не полагайтесь на безопасность оружия, чтобы защитить вас от небезопасного оружия.
умение обращаться. Предохранитель — это всего лишь механическое устройство, а не замена
за использование здравого смысла и соблюдение правил безопасности при обращении с оружием.
Учитесь у
профессионал, как обращаться, загружать, разгружать, эксплуатировать, стрелять и ухаживать
для вашего пистолета. НИКОГДА НЕ ИГРАЙТЕ со своим огнестрельным оружием. Это не игрушка и может
быть очень опасным оружием, которое может привести к серьезным травмам
или смерть. Всегда держите пистолет направленным в безопасном направлении. Удовольствие
каждое ружье как будто заряжено…все время. НИКОГДА не указывайте
огнестрельное оружие (заряженное или незаряженное) против любого человека или чего-либо, что вам не
намерен стрелять. Всегда держите ружье незаряженным, пока оно не будет готово к использованию.
и убедитесь, что он разряжен перед очисткой. Держи палец
ОТКЛЮЧАЙТЕ спусковой крючок до тех пор, пока вы не прицелитесь в цель и
готов стрелять. Научитесь удерживать палец вне спускового крючка
сторожить. Никогда не полагайтесь на безопасность оружия, чтобы защитить вас от небезопасного оружия.
умение обращаться. Предохранитель — это всего лишь механическое устройство, а не замена
за использование здравого смысла и соблюдение правил безопасности при обращении с оружием.
5
СОБЛЮДЕНИЕ ЭТИХ ПРАВИЛ БЕЗОПАСНОСТИ МОЖЕТ СОХРАНИТЬ ВАШУ ЖИЗНЬ ИЛИ ЖИЗНЬ ДРУГИХ
КТО-ТО ДРУГОЙНикогда не оставляйте оружие без присмотра или там, где оно может упасть.
Храните оружие и боеприпасы отдельно от детей.
Убедитесь, что они надежно закреплены, чтобы необученные люди
и детям будет отказано в доступе к вашему оружию и боеприпасам.
Проверяйте работу и безопасность своего оружия только на стрельбище
пока огнестрельное оружие направлено в безопасном направлении. Знай свою цель
и что дальше. Спросите себя, во что попадет снаряд, если он
не попадает в цель или проходит через намеченную цель. Запомнить,
выпущенный снаряд может пролететь более мили (включая рикошет)
и способен проходить сквозь стены. Никогда не стреляйте в твердую плоскость
поверхностей или воды. . .выстрел рикошетом. Оружие и алкоголь или наркотики
не смешивать. Не принимайте алкогольные напитки и/или наркотики до или
во время съемок. Никогда не тяните пистолет к себе за
морда. Не лезьте на дерево, не пересекайте канаву или забор с грузом. пистолет. Всегда разряжайте ружья перед тем, как войти в дом, машину, грузовик, лодку,
RV, лагерь или любое здание. При получении оружия всегда открывайте
действия и убедитесь, что патронник не заряжен. НИКОГДА не принимайте
Кто-нибудь скажет, что ружье разряжено или пусто. Вам следует
немедленно откройте действие и убедитесь, что оно выгружено.
Будьте осторожны со ВСЕМИ боеприпасами. Даже холостые патроны смертельны при
с близкого расстояния из-за дульного выстрела. Убедитесь, что ваши боеприпасы
новый и в хорошем чистом состоянии. НЕ смазывайте боеприпасы маслом или консистентной смазкой, т.к.
это может повредить капсюль картриджа. ЗАПРЕЩАЕТСЯ вмешиваться или изменять
штатные заводские патроны. Держите огнестрельное оружие незаряженным, когда не
фактически в использовании. Заряжайте ружье только на стрельбище, готовясь к стрельбе.
стреляйте и разряжайте его, прежде чем покинуть полигон. Никогда не клади руку
над дулом пистолета. ВСЕГДА НОСИТЕ защитные очки для стрельбы
и защита органов слуха при использовании огнестрельного оружия.
пистолет. Всегда разряжайте ружья перед тем, как войти в дом, машину, грузовик, лодку,
RV, лагерь или любое здание. При получении оружия всегда открывайте
действия и убедитесь, что патронник не заряжен. НИКОГДА не принимайте
Кто-нибудь скажет, что ружье разряжено или пусто. Вам следует
немедленно откройте действие и убедитесь, что оно выгружено.
Будьте осторожны со ВСЕМИ боеприпасами. Даже холостые патроны смертельны при
с близкого расстояния из-за дульного выстрела. Убедитесь, что ваши боеприпасы
новый и в хорошем чистом состоянии. НЕ смазывайте боеприпасы маслом или консистентной смазкой, т.к.
это может повредить капсюль картриджа. ЗАПРЕЩАЕТСЯ вмешиваться или изменять
штатные заводские патроны. Держите огнестрельное оружие незаряженным, когда не
фактически в использовании. Заряжайте ружье только на стрельбище, готовясь к стрельбе.
стреляйте и разряжайте его, прежде чем покинуть полигон. Никогда не клади руку
над дулом пистолета. ВСЕГДА НОСИТЕ защитные очки для стрельбы
и защита органов слуха при использовании огнестрельного оружия.
6
СОБЛЮДЕНИЕ ЭТИХ ПРАВИЛ БЕЗОПАСНОСТИ МОЖЕТ СОХРАНИТЬ ВАШУ ЖИЗНЬ ИЛИ ЖИЗНЬ ДРУГИХ
КТО-ТО ДРУГОЙ Зрители должны находиться не менее чем в 10 футах позади и в стороне.
из стрелка, пока стрелок заряжает, стреляет и
разгрузка. Зрители должны носить средства защиты слуха и зрения.
потеря, и они должны избегать отвлекать тех, кто стреляет в их
огнестрельное оружие. Никогда не бросайте пистолет. Если вы его уроните, выгрузите его и
проверьте его правильность работы, прежде чем использовать его снова. НЕ изменяйте или
модифицировать свое оружие. Не пытайтесь изменить усилие спускового крючка вашего оружия, потому что
это может повлиять на зацепление шептала и тем самым вызвать случайное срабатывание.
ЗАПРЕЩАЕТСЯ снимать какие-либо внутренние предохранители или предохранительные устройства на огнестрельном оружии.
Механизмы безопасности предназначены для того, чтобы защитить вас от травм или
смерть. Правильно ухаживайте за своим оружием и держите его в чистоте. Держите его в сухом
место вдали от других металлов и воды. Если на вашем пистолете видны признаки
коррозия или неправильная эксплуатация, обратитесь за обслуживанием к компетентному
оружейник. Держите дуло направленным в безопасном направлении при заряжании
и разрядить ружье. Никогда не направляйте морду ни на кого. Если пистолет
не срабатывает при нажатии на спусковой крючок, держите его направленным вниз
дальность до цели в течение 60 секунд. Иногда медленное воспламенение капсюля
может привести к задержке полного воспламенения патрона. Если после 60
секунд, патрон не стреляет, держите дуло в безопасном положении
направлении и избегайте воздействия на затвор, поскольку вы пытаетесь
разрядите свое огнестрельное оружие. Никогда не допускайте попадания воды, снега, грязи или других материалов
войти в бочку. Всегда следите за тем, чтобы на стволе не было
препятствие. Поскольку многие обозначения боеприпасов звучат одинаково,
убедитесь, что вы используете только правильные боеприпасы для своего оружия. Делать
ЗАПРЕЩАЕТСЯ использовать боеприпасы неправильного размера или силы в своем
огнестрельное оружие.
Если на вашем пистолете видны признаки
коррозия или неправильная эксплуатация, обратитесь за обслуживанием к компетентному
оружейник. Держите дуло направленным в безопасном направлении при заряжании
и разрядить ружье. Никогда не направляйте морду ни на кого. Если пистолет
не срабатывает при нажатии на спусковой крючок, держите его направленным вниз
дальность до цели в течение 60 секунд. Иногда медленное воспламенение капсюля
может привести к задержке полного воспламенения патрона. Если после 60
секунд, патрон не стреляет, держите дуло в безопасном положении
направлении и избегайте воздействия на затвор, поскольку вы пытаетесь
разрядите свое огнестрельное оружие. Никогда не допускайте попадания воды, снега, грязи или других материалов
войти в бочку. Всегда следите за тем, чтобы на стволе не было
препятствие. Поскольку многие обозначения боеприпасов звучат одинаково,
убедитесь, что вы используете только правильные боеприпасы для своего оружия. Делать
ЗАПРЕЩАЕТСЯ использовать боеприпасы неправильного размера или силы в своем
огнестрельное оружие. Научите детей: 1. НЕ прикасаться к оружию, пулям, патронам,
снаряды, сигнальные ракеты или любое взрывное устройство и 2. Если они обнаружат
такой опасный предмет или оружие, они должны немедленно сообщить о его
своим родителям, школьному учителю или полиции. Помните
огнестрельное оружие способно лишить вас жизни или жизни
кто-нибудь другой. Будьте осторожны с огнестрельным оружием, несчастный случай почти
всегда является результатом несоблюдения основных правил безопасности.
Научите детей: 1. НЕ прикасаться к оружию, пулям, патронам,
снаряды, сигнальные ракеты или любое взрывное устройство и 2. Если они обнаружат
такой опасный предмет или оружие, они должны немедленно сообщить о его
своим родителям, школьному учителю или полиции. Помните
огнестрельное оружие способно лишить вас жизни или жизни
кто-нибудь другой. Будьте осторожны с огнестрельным оружием, несчастный случай почти
всегда является результатом несоблюдения основных правил безопасности.
7
Используйте замок безопасности, прилагаемый к вашему ружьюЭто ружье
поставляется с замком безопасности, чтобы помочь в защите от
несанкционированное использование. См. рисунок A. Ружье должно быть разряжено, когда
замок безопасности находится на месте на ружье. ПРЕДУПРЕЖДЕНИЕ: Использование
замок безопасности не может заменить защиту вашего огнестрельного оружия и
боеприпасы в отдельных, запертых местах. Замок безопасности
работает следующим образом: 1. Чтобы открыть триггерный замок, вставьте ключ в
пазы стопорной гайки спускового крючка и отвинтите ее. против часовой стрелки. См. рис. 1 и 2. Снимите блокировку курка.
Винт.
против часовой стрелки. См. рис. 1 и 2. Снимите блокировку курка.
Винт.
Рисунок A
Рисунок 1
Рисунок 2
ружье поместите замок спускового крючка вокруг спусковой скобы. См. рисунок 3. 3. Наверните стопорную гайку спускового крючка на стопорный винт спускового крючка. (по часовой стрелке) и затяните гайку ключом. НЕ ЗАТЯГИВАЙТЕ ЗАТЯГИВАЙТЕ ГАЙКУ СЛИШКОМ ЗАТЯЖНО, ИЛИ ВЫ МОЖЕТЕ ПОВРЕДИТЬ СТОПОРНЫЙ ВИНТ. См. рис. 4. 4. Чтобы снять блокировку курка, отвинтите стопорную гайку курка с помощью ключ и отсоедините стопорный винт спускового крючка. См. рис. 5. Рис. 3
Рисунок 4
ПРЕДУПРЕЖДЕНИЕ Всегда храните ключ от защитного замка: (1) в
отдельное расположение подальше от дробовика; (2) в безопасном месте
неизвестно и недоступно другим, особенно детям. Будь то
вы защищаете свое огнестрельное оружие Figure 5 с помощью прилагаемой защиты
замок, замок пистолета или сейф или какой-либо другой механизм, это ВАШЕ
ОТВЕТСТВЕННОСТЬ за хранение огнестрельного оружия таким образом, чтобы
дети и другие посторонние лица не могут получить доступ к
их.
8
ИСПОЛЬЗУЕМОЕ ОГНЕСТРЕЛЬНОЕ ОРУЖИЕ — огнестрельное оружие иногда модифицируют, чтобы оно работало. неправильно, или части могут быть удалены, потеряны или заменены неправильные детали. Если у вас есть европейско-американская оружейная корпорация. огнестрельное оружие как бывшее в употреблении, ПЕРЕД ЕГО ИСПОЛЬЗОВАНИЕМ вы должны отнести его к хорошему оружейник, знающий огнестрельное оружие European American Armory Corp., и пусть он осмотрит его. НАПИСАТЬ Европейско-американскую оружейную корпорацию. относительно любых предметов или обстоятельств, которые вы не понимаете и которые могут иметь отношение к вашей безопасности и эксплуатации вашего огнестрельного оружия по адресу: European American Armory Corp., 402 Richard Road, Rockledge, Флорида 32955, тел.: (321) 639-4842.
ПРЕДУПРЕЖДЕНИЕ (ВОЗМОЖНЫЕ ПРЕПЯТСТВИЯ) ПЕРЕД ЗАРЯЖЕНИЕМ или выстрелом
ружье, осмотрите канал ствола и патронник, чтобы убедиться, что они
чисты и свободны от каких-либо препятствий. Даже толстый слой масла,
жир, снег или вода могут привести к повреждению ружья и
ранение стрелка и/или лиц, находящихся рядом с огнестрельным оружием. Осечка или
необычный звук при стрельбе является сигналом к прекращению стрельбы и
осмотрите патронник и канал ствола. Если есть препятствие, устраните
препятствия и очистите канал ствола и патронник перед выстрелом. Любой
закупорка отверстия, даже если оно заблокировано лишь частично, может вызвать
пистолет взорвется при выстреле или может привести к повреждению оружия, например
как выпуклая бочка. Во избежание ранения или смерти стрелка или
прохожие, проверяйте ствол перед выстрелом или если выстрел не
звук нормальный.
Осечка или
необычный звук при стрельбе является сигналом к прекращению стрельбы и
осмотрите патронник и канал ствола. Если есть препятствие, устраните
препятствия и очистите канал ствола и патронник перед выстрелом. Любой
закупорка отверстия, даже если оно заблокировано лишь частично, может вызвать
пистолет взорвется при выстреле или может привести к повреждению оружия, например
как выпуклая бочка. Во избежание ранения или смерти стрелка или
прохожие, проверяйте ствол перед выстрелом или если выстрел не
звук нормальный.
ПРЕДУПРЕЖДЕНИЕ Всегда направляйте дуло в безопасном направлении! Никогда
пытаться заряжать или разряжать любое огнестрельное оружие внутри транспортного средства, здания или
иное замкнутое пространство (кроме правильно устроенного стрелкового
диапазон). Закрытые помещения часто не предлагают полностью безопасных
направление, в котором нужно направить огнестрельное оружие. Если случайный выброс
происходит, существует большой риск смерти, телесных повреждений или повреждения имущества
наносить ущерб. Перед загрузкой всегда очищайте все, смазку и масло от
отверстие и патронник и убедитесь, что в
бочка. Любой посторонний предмет в стволе может привести к
вздутие или разрыв ствола или другие повреждения огнестрельного оружия и могут
нанести серьезную травму стрелку или другим лицам.
Перед загрузкой всегда очищайте все, смазку и масло от
отверстие и патронник и убедитесь, что в
бочка. Любой посторонний предмет в стволе может привести к
вздутие или разрыв ствола или другие повреждения огнестрельного оружия и могут
нанести серьезную травму стрелку или другим лицам.
ПРЕДУПРЕЖДЕНИЕЕсли есть основания подозревать, что снаряд блокируя ствол, немедленно разрядите ружье (см. Инструкции по разгрузке) и проверьте патронник и канал ствола. пыж, выстрел или какой-либо другой предмет может быть заложен на некотором расстоянии вниз по ствол там, где его не видно. Проверьте отверстие с помощью чистящий стержень для прохождения через ствол. Если блокировка находится в DO
9
НЕ ПЫТАЙТЕСЬ ВЫСТРЕЛИВАТЬ ЕГО ДРУГИМ ПАТРОНОМ ИЛИ УДАРИТЬ ЕГО
УДАЛИТЬ ХОЛОСТЫМ ПАТРОНОМ ИЛИ ПАТРОНОМ, ИЗ КОТОРОГО БЫЛ ВЫСТРЕЛ
БЫЛ УДАЛЕН. ТАКАЯ ТЕХНИКА (МЕТОДЫ) МОГУТ СОЗДАТЬ ЧРЕЗМЕРНОЕ ДАВЛЕНИЕ,
ПОВРЕДИТЬ ОРУЖИЕ И ПРИЧИНИТЬ СЕРЬЕЗНУЮ ТРАВМУ. Если
засор можно удалить с помощью шомпола, прочистить все несгоревшие
пороховые зерна из канала ствола, патронников и механизма перед возобновлением
стрельба. Если засор не удается устранить, постукивая по нему
чистящий стержень, отнесите огнестрельное оружие оружейнику.
Если засор не удается устранить, постукивая по нему
чистящий стержень, отнесите огнестрельное оружие оружейнику.
ПРЕДУПРЕЖДЕНИЕ (ЗАЩИТА ГЛАЗ И УШЕЙ) и защита ушей обязательна во все времена. Стрелки и зрители должны носить стрелковые очки и средства защиты органов слуха, чтобы предотвращения травм глаз и потери слуха. Нарушение зрения и слуха может произойти только с одним моментом отсутствия защиты.
ПРЕДУПРЕЖДЕНИЕ (Механические неисправности) НЕМЕДЛЕННО ПРЕКРАТИТЕ СЪЕМКУ
И РАЗРЯДИТЕ СВОЕ ОГНЕСТРЕЛЬНОЕ ОРУЖИЕ, если в вашем огнестрельном оружии:
неисправность; связывание или остановка; плеваться порошком/газом; проколотый
патронный капсюль; вздутая или разорванная гильза; или звук
при стрельбе звучит не совсем правильно. НЕ делайте еще один выстрел, но
разрядите свое огнестрельное оружие и отнесите его и боеприпасы квалифицированному специалисту.
оружейника или отправить огнестрельное оружие обратно в European American Armory Corp.
на экспертизу. Не думайте, что дробовик пуст только
потому что ты проверил камеру. Также необходимо проверить отверстие на предмет
любой произведенный выстрел или пыж застрял внутри ствола.
Также необходимо проверить отверстие на предмет
любой произведенный выстрел или пыж застрял внутри ствола.
ОСНОВНЫЕ ЧАСТИ ВАШЕГО РУЖЬЯ ВНЕШНИЕ ДЕТАЛИ УПРАВЛЕНИЯ: спусковой крючок Безопасность: предохранительный механизм ружья обеспечивает защиту от случайного и непреднамеренного разряда при нормальном использовании при правильном включении и в хорошем рабочем состоянии. Безопасность механизм не является заменой соблюдения правил безопасного обращения с оружием умение обращаться. Не отключайте предохранитель, если вы не готовы стрелять дробовик. УБЕДИТЕСЬ, ЧТО ВЫ ПОНИМАЕТЕ, КАК РАБОТАЕТ БЕЗОПАСНОСТЬ, ПЕРЕД ВЫ ЗАГРУЗИТЕ ИЛИ ИСПОЛЬЗУЕТЕ РУЖЬЕ. Механизм безопасности представляет собой хвостовик. предохранитель, расположенный за верхним рычагом. См. рис. 6. Чтобы разместить безопасность на
Рисунок 6
10
или безопасное положение, потяните кнопку до упора назад. Когда
во включенном или безопасном положении предохранитель блокирует движение назад
триггера(ов). Чтобы перевести предохранитель в выключенное или огневое положение,
сдвиньте ползунок вперед до упора. Когда в выключенном состоянии или огонь
положение, за ползунковой кнопкой видна красная точка. У вас есть
Теперь предохранитель отключен, и ружье находится в состоянии готовности к стрельбе.
должность. См. рис. 7.
ПРЕДУПРЕЖДЕНИЕ
Рис. 7
Не прикасайтесь к спусковому крючку во время перемещения предохранителя. Когда при работе с предохранителем убедитесь, что вы нажимаете или тянете его до упора. степень его движения в желаемом направлении, либо полностью или полностью выключен. Полубезопасный или частично безопасный НЕБЕЗОПАСНЫЙ.
Верхний рычаг: верхний рычаг расположен прямо перед предохранителем.
механизм. См. рис. 8. Верхний рычаг используется для взлома
действия по зарядке и/или разрядке ружья и сборке Рис. 8
и разобрать ружье. Чтобы открыть действие, полностью нажмите
верхний рычаг вправо. См. рис. 9.. Триггер(ы): Больше и
под ружья от European American Armory Corp. доступны в
либо конфигурация с двойным триггером, либо одиночный триггер
конфигурация. См. рис. 10 и рис. 11. Ружья с двойным спусковым крючком
сделаны по образцу классических вертикальных и нижних ружей с двумя
триггеры. Передний спусковой крючок всегда стреляет из нижнего ствола.
задний спусковой крючок всегда стреляет из верхнего ствола. Нажатие любого триггера
с предохранителем в выключенном или огневом положении производит выстрел из дробовика.
Рисунок 9
Рисунок 10
ПРЕДУПРЕЖДЕНИЕ
Никогда не нажимайте на спусковой крючок до тех пор, пока вы действительно не будете готовы к стрельбе. Держите пальцы подальше от спускового крючка (спусковых крючков) и за пределы спускового крючка. охраняйте, пока не будете готовы нажать на курок и выстрелить из ружья. При открытии и закрытии затвора, а также при включении или отключая предохранительный механизм, убедитесь, что ваши пальцы вне спусковой скобы и вдали от спускового крючка(ов).
Рисунок 11
11
ПРЕДУПРЕЖДЕНИЕ Никогда не стреляйте из двуспускового ружья двумя пальцами
потому что вы можете непреднамеренно нажать на оба триггера одновременно
что привело к почти одновременному выстрелу из обоих стволов. Используй свой
указательным пальцем, чтобы нажать на один из спусковых крючков, чтобы произвести первый выстрел.
После этого выстрела отпустите спусковой крючок и переместите указатель
палец к другому спусковому крючку, чтобы произвести второй выстрел. Одиночный триггер
Ружья ИЖ37 имеют один спусковой крючок, который используется для выстрела из каждой
бочка. Как только спусковой крючок нажат для выстрела из первого ствола,
отпустите курок. После этого второй ствол готов к выстрелу.
снова нажать на спусковой крючок. Эжекторы: ружья ИЖ37
оборудованы эжекторами, настроенными на заводе на
автоматический выброс отстрелянных патронов при открытии затвора.
Однако пользователь может отрегулировать эжекторы, чтобы отключить
функция автоматического выброса. Когда эжекторы установлены в
отключенное положение, выпущенные снаряды не будут автоматически выбрасываться
из стволов при открытии затвора. Эжекторы
управляется установочными винтами, расположенными на изогнутой части спереди
приемника. Правый установочный винт управляет выбросом
снаряды снизу Рисунок 12 Рисунок 13 ствол. См. рис. 12.
Левый установочный винт управляет выбросом гильз из ствола.
верхняя бочка. См. рис. 13. Установочные винты для выталкивателей
установлен на заводе для автоматического выброса гильз при
действие открыто. При автоматической настройке слоты в
установить Рисунок 15 Рисунок 14 винты ориентированы в вертикальном положении
идущие перпендикулярно длине стволов. См. вертикальный
расположение слотов, как показано на рисунках 14 и 15. Чтобы отключить
автоматические эжекторы, откройте затвор и поверните установочные винты
по часовой стрелке так, чтобы пазы для установочных винтов находились в горизонтальном положении.
положение параллельно длине стволов. См. рисунки 16 и
17.
Рисунок 16
Рисунок 17
12
Чтобы вернуть ружье в режим автоматического выброса, просто
поверните установочные винты против часовой стрелки на один оборот в исходное положение
положение с установочным винтом Рис. 18 Рис. 19 пазы ориентированы
вертикальное положение перпендикулярно длине стволов. Видеть
Рисунки 18 и 19.
ПРЕДУПРЕЖДЕНИЕ. Никогда не регулируйте и не отключайте установочные винты выталкивателя, когда дробовик заряжен. Регулируйте установочные винты только тогда, когда ружье полностью разгружен.
ПРЕДУПРЕЖДЕНИЕ Никогда не выкручивайте установочные винты выбрасывателя из ружья. эжекторы входят в состав системы поддержки головки гильзы. Стрельба ружье со снятыми или смещенными эжекторами может вызвать разрыв головки корпуса, что привело к серьезной травме.
Переключатель ствола (только для односпусковых ружей): как
в заводской комплектации односпусковое ружье модели ИЖ37
настроен на выстрел из нижнего ствола при первом выстреле, а затем
верхний ствол на втором выстреле. Как только дробовик заряжен, вы можете
вручную настроить дробовик так, чтобы сначала стрелял верхний ствол. Выбирать
верхний ствол, с включенным предохранителем, точка
дульный срез в безопасном направлении и нажмите на спусковой крючок вперед из
сзади, пока не почувствуете и не услышите щелчок. Теперь вы установили
дробовик для стрельбы из верхнего ствола при первом выстреле и из нижнего
ствол на втором выстреле. Примечание. Как только вы откроете действие с
верхний рычаг, дробовик автоматически возвращается в исходное положение
конфигурация стрельбы из нижнего ствола в первую очередь.
ПРЕДУПРЕЖДЕНИЕ. При нажатии курка вперед для выбора верхний ствол, убедитесь, что предохранитель включен, а дуло направлено внутрь безопасное направление. НИКОГДА не используйте селектор ствола, когда предохранитель в выключенном или огневом положении. После того, как вы использовали триггер для выберите верхний ствол, уберите палец изнутри спускового крючка охраняйте, пока не будете готовы стрелять.
13
Цевье: Цевье представляет собой деревянную рукоятку, которая крепится
к узлу ствола и фиксирует узел ствола в
получатель. См. рис. 20. На цевье имеется механическая защелка,
используется для блокировки и разблокировки цевья к стволу в сборе. Никогда
попытаться выстрелить из дробовика, не зафиксировав цевье
правильное положение на стволе.
Рисунок 20
Серийный номер: Серийный номер ружья находится на верхней части ствольной коробки под верхним рычагом.
ИНСТРУКЦИЯ ПО СБОРКЕ РУЖЬЯ: ВНИМАНИЕ! собрать дробовик, вы ДОЛЖНЫ (1) смотреть вниз стволы из казенную часть к дульному концу, чтобы убедиться, что нет боеприпасы в патронниках или препятствия в стволах, и (2) убедитесь, что предохранитель находится в безопасном положении. Ваш дробовик был поставляется со стволом в сборе (ствол и цевье), отсоединенным от ресивер в сборе. Чтобы собрать дробовик, следуйте этим Инструкции: 1) Снимите полиэтиленовые пакеты или упаковку, закрывающую части. Затем снимите цевье со ствола, потянув наружу за механический рычаг в нижней средней части цевья и наклон цевья наружу и в сторону от ствола. См. рис. 21. 2) Теперь нажмите верхний рычаг ствольной коробки вправо и удерживайте с одной стороны. Не нажимайте на верхний рычаг. См. рис. 22.
Рисунок 21
3) Удерживая стволы другой рукой, включите нижний фиксатор
выступ расположен на нижней части стволов со ствольной коробкой. Полукруглый вырез на нижнем фиксирующем выступе соответствует
круглый стержень на Рисунке 22, передняя нижняя часть паза в
получатель. Вставив стволы в ствольную коробку, поверните
стволы вверх в заблокированное положение. Верхний рычаг должен двигаться назад
в центр после того, как стволы зафиксируются на месте. Обратите внимание
верхний рычаг может не двигаться полностью в центр на новом ружье
до тех пор, пока ружье не будет прикручено. Верхний рычаг должен перемещаться не менее чем
три
14
четверти полного пути. См. рисунок 23 (а, б, в).
Рисунок 23a
Рисунок 23b
Рисунок 23c
4) Теперь, когда стволы зафиксированы в ствольной коробке, вы
необходимо снова прикрепить цевье к стволам. Поместите цевье на
передний изгиб ствольной коробки (угол между цевьем и
стволы должны быть около 20 градусов). Держите контакт между передним
и ресивер и качающим движением вверх заблокируйте Рисунок 24
цевье в стволы. Хотя может быть необходимо нажать
защелку в окончательное плоское положение, никогда не применяйте силу
должность. См. рис. 24.
ПРЕДУПРЕЖДЕНИЕ Никогда не стреляйте из ружья, если стволы не заперты. в положение на ствольной коробке, а цевье зафиксировано в ресивер и стволы.5) Убедитесь, что предохранитель спускового крючка включается после завершения сборки (красная точка покрыта). См. рисунок 25.
ЗАРЯДКА И РАЗРЯДКА РУЖЬЯ
Рис. 25
ВНИМАНИЕ! зафиксированы в положении на ствольной коробке, а цевье заблокировано в ствольную коробку и стволы. ПРЕДУПРЕЖДЕНИЕ Убедитесь, что дробовик направлен в безопасном направлении, а предохранитель спускового крючка постоянно задействованы при погрузке и разгрузке. НИКОГДА не позволяйте своему пальцами или другими предметами, чтобы коснуться спускового крючка во время заряжания или разгрузка. ПРЕДУПРЕЖДЕНИЕ Не выключайте предохранитель блока спускового крючка. до тех пор, пока вы не будете готовы стрелять и безопасно навести пистолет вниз
15
ПРЕДУПРЕЖДЕНИЕ Не заряжайте ружье, пока не будете готовы к стрельбе. К
НАГРУЗКА: 1) Убедитесь, что боеприпасы, которые вы используете, являются правильными. размер и калибр. Ствол маркируется калибром и патронником
размер. Стволы с маркировкой 3 дюйма 76 мм будут использовать только 2 3/4 дюйма (70 мм) или 3 дюйма.
(76 мм) и не должны использоваться со снарядами 3 1/2 дюйма (89 мм).
Стволы с маркировкой 2 3/4 дюйма (70 мм) будут использовать только 2 3/4 дюйма (70 мм) и
нельзя использовать с гильзами диаметром 3 дюйма (76 мм) или 3 1/2 дюйма (89 мм). 2)
Визуально осмотрите патронники и стволы на наличие препятствий с помощью
открыть ружье, нажав на запорный рычаг и
опрокидывание стволов вниз. Патронники и стволы должны быть свободны
масла или смазки или препятствий. См. рис. 26.
Рисунок 26
3) После проверки загрузите дробовые гильзы правильного типа в
камеры, убедившись, что гильзы находятся на одном уровне с
экстрактор. См. рисунки Рисунок 28 Рисунок 27 27 и 28). Затем закрыть
действие, поворачивая стволы вверх, пока они не зафиксируются в
получатель. Запорная защелка должна переместиться в полностью запертое положение.
должность. См. рис. 29. Если рычаг блокировки не перемещается в
полностью заблокированное положение. (см. рис. 30) снова открыть затвор
и проверьте, полностью ли вошла гильза в патронник.
Если оболочки полностью установлены, снова закройте Рисунок 29.Рисунок 30
Действие. Если гильзы вставлены не полностью, проверьте
камеры на засорение и проверить обечайки на деформацию и
повторите попытку закрытия действия еще раз. Если снаряды правильные
типа и патронники не забиты и гильзы не
деформирован, и верхний рычаг не перемещается в полностью заблокированное положение.
положение, разрядите пистолет (следуйте инструкциям по разгрузке) и
грамотный оружейник посмотрите на пистолет или верните пистолет европейцу
American Armory Corp. для проверки. Не форсируйте дробовик
закрыто.
16
ДЛЯ РАЗРЯДКИ: 1) Откройте ружье, нажав на верхний рычаг
ствольную коробку до упора вправо и опрокидывание стволов вниз.
См. рис. 31. 2) Извлеките оболочку(и) из камеры(ей),
хватая их за края и вытягивая из камер. Видеть
Рисунок 32. 3) Визуально проверьте стволы, чтобы убедиться, что они
свободен от препятствий. См. Рисунок 33.
Рисунок 31
Рисунок 32
СТРЕЛЬБА ИЗ РУЖЬЯ ВНИМАНИЕ! патронник, пока не будете готовы выстрелить из дробовика.
Рисунок 33
ПРЕДУПРЕЖДЕНИЕ Не отключайте предохранитель спускового крючка, пока дробовик направлен в безопасном направлении, и вы готовы Огонь. ПРЕДУПРЕЖДЕНИЕ Если патрон не стреляет, а спусковой крючок нажат и предохранитель спускового крючка отключен, Стоп! Затем сделайте следующее: а) Убедитесь, что пистолет направлен в сейф. направлении b) Включите предохранитель спускового крючка c) Подождите 60 секунд d) Следуйте приведенным выше инструкциям по разрядке ружья.
ВНИМАНИЕ! Если во время стрельбы звук любого патрона заметно
тише или громче, чем выстрелил предыдущий патрон, Стоп! Тогда сделайте
следующее: а) Убедитесь, что пистолет направлен в безопасном направлении.
б) Включите предохранитель спускового крючка. в) Следуйте приведенным выше инструкциям. для разрядки ружья и d) с разряженным ружьем визуально
осмотрите ружье на предмет засорения или повреждения ствола. Затем осмотрите
ствольную коробку ружья на наличие повреждений, прежде чем продолжить.
17
ПРОЧТИТЕ, ПОНИМИТЕ И СОБЛЮДАЙТЕ ВСЕ ПРЕДУПРЕЖДЕНИЯ И ИНСТРУКЦИИ В
В ЭТОМ РУКОВОДСТВЕ ПЕРЕД СТРЕЛЬБОЙ. ДЛЯ СТРЕЛЯ ИЗ РУЖЬЯ: 1) Сделать
Убедитесь, что вы носите защиту для глаз и ушей. 2) Убедитесь, что
ружье направлено в безопасном направлении спусковым механизмом
включена безопасность. См. рис. 34. 3) Держите пальцы подальше от
спусковой крючок(и), переведите предохранитель спускового крючка в положение выключения или выстрелите
положение с красной точкой. См. рис. 35. 4) С прикладом
приклад плотно прижат к плечу, а ствол направлен
к намеченной цели, поместите указательный палец на спусковой крючок.
курок. Дробовик выстрелит, когда вы нажмете или нажмете на спусковой крючок.
с необходимой силой. См. рис. 36. 5) Если вы хотите запустить
второго выстрела из другого ствола необходимо: а) В случае
Односпусковое ружье модели ИЖ37, отпустите курок и затем
потяните или сожмите его снова. Затем выстрелит второй ствол.Рисунок
34
Рисунок 35
Рисунок 36
б) В случае двухспускового ружья модели ИЖ37 переместите палец к другому курку и потяните или сожмите этот курок. Затем выстрелит второй ствол.
ПРЕДУПРЕЖДЕНИЕ Как только вы прекратите стрелять из дробовика, немедленно потяните предохранитель спускового крючка обратно во включенное или безопасное положение. должность. Затем следуйте приведенным выше инструкциям по разгрузке, пока направляя дуло в безопасном направлении. НАПРАВЛЕНИЯ ДЛЯ РАЗБОРКА РУЖЬЯ: 1) Убедитесь, что ружье разряжено. (СЛЕДУЙТЕ ИНСТРУКЦИЯМ ПО РАЗГРУЗКЕ) и указал в безопасном направлении. 2) Убедитесь, что предохранитель спускового крючка находится во включенном или безопасном положении. должность. См. Рисунок 37. Рисунок 37
18
3) Убедитесь, что узел ствола зафиксирован в ствольной коробке и
акция закрылась. Верхний рычаг теперь должен быть в центре
должность. Снимите цевье со ствола, вытянув
механический рычаг в нижней центральной части цевья и снятие
цевье в качающем движении вниз наружу. См. рис. 38. Никогда
снимите цевье со ствола и ствольной коробки. 4) С бочками
и ствольную коробку, поддерживая ее одной рукой, полностью нажмите на верхний рычаг, чтобы
вправо и наклоните узел ствола вниз, пока он не выйдет из зацепления.
получатель. Проверьте стволы на наличие повреждений и препятствий после
удаление. См. рис. 39..
Рисунок 38
Рисунок 39
ПРЕДУПРЕЖДЕНИЕ ПРОИЗВОДИТЕЛЯ.
работайте с оригинальными деталями, как задумано. Это ваша обязанность
убедитесь, что все детали, которые вы покупаете, правильно установлены и что
ни замены, ни оригиналы не переделываются и не изменяются. Ваш пистолет
представляет собой сложный инструмент со многими частями, которые должны правильно относиться к
другие детали для безопасной и точной работы. Собираем пистолет
неправильные или модифицированные детали могут привести к повреждению пистолета, или
телесные повреждения или смерть для вас или других лиц. Всегда позволяйте квалифицированному
работа оружейника над вашим ружьем или, по крайней мере, проверка невыполненных работ
оружейником. Безопасность огнестрельного оружия — ваша главная задача. ПИСТОЛЕТ
ВЛАДЕЛЕЦ ДОЛЖЕН ПРИНЯТЬ ПОЛНУЮ ОТВЕТСТВЕННОСТЬ ЗА ПРАВИЛЬНУЮ СБОРКУ
И ФУНКЦИОНИРОВАНИЕ ОРУЖИЯ ПОСЛЕ ЛЮБОЙ РАЗБОРКИ ИЛИ ЗАМЕНЫ
ЧАСТЕЙ.
ДЛЯ ЧИСТКИ И ОБСЛУЖИВАНИЯ ВАШЕГО РУЖЬЯ: A) Следуйте процедурам для
разряжать ружье, как описано выше. B) Следуйте процедурам для
разборка ружья, как описано выше. В) Необходимые материалы
для чистки этого ружья: шомпол, ватные тампоны,
щетка из латуни, порошковый растворитель, небольшая мягкая щетка и хороший
качественное оружейное масло. ПРЕДУПРЕЖДЕНИЕ: Свинец или соединения свинца известны
Штат Калифорния вызывает рак, врожденные дефекты, репродуктивную
токсичность и другие серьезные телесные повреждения. Те, кто чистит
огнестрельное оружие должны принимать защитные меры, чтобы избежать контакта или
воздействие таких химических веществ. D) Использование щетки для отверстий с порошком
растворителя, протрите внутреннюю часть стволов и патронников, чтобы удалить
любой порох и остатки свинца в канале ствола. Всегда и в последнюю очередь мазок
внутреннюю часть стволов ватным тампоном с оружейным маслом для покрытия
и защитить канал ствола и камеры от ржавчины и коррозии.
19
E) Любое пороховое загрязнение на огневой стенке, стволе и т. д. может быть
легко чистится щеткой и порошковым растворителем. F) Все внешние
поверхности должны быть протерты легким слоем ржавчины
профилактический. Избегайте использования слишком большого количества масла в виде порошка, пыли или других
посторонние предметы могут попасть в масло. Это может привести к
застывшие отложения, которые могут помешать безопасному и надежному
эксплуатации дробовика. G) БУДЬТЕ ОСТОРОЖНЫ при использовании растворителей в
чистка канала ствола оружия. Длительный или чрезмерный контакт с растворителями может
повредить пушки воронением и отделкой. Обязательно удалите все лишнее
растворителем, а затем смажьте маслом. H) Оружие должно быть очищено
после стрельбы. Кроме того, внешние части следует протирать
промасленная ткань после обработки. Легкая нефть – это обычно все, что
необходима в чистке, если из ружья не стреляли или стреляли очень
маленький. Если пистолет используется часто, его необходимо очищать после каждого
стрельбы и регулярно обслуживается профессиональным оружейником. Если
пистолет хранился, удалите все лишнее масло и/или смазку перед
стрельба. Всегда проверяйте, нет ли чистящего пластыря или другого
перед выстрелом в канале ствола или патронниках остается препятствие.
ОПАСНО БОЕПРИПАСЫ ПРЕДУПРЕЖДЕНИЕОгнестрельное оружие может быть повреждено и серьезно
может привести к телесным повреждениям или смерти стрелка или прохожих.
от любого состояния, которое способствует возникновению чрезмерного
давление или неконтролируемый выброс газа в огнестрельном оружии. Такой
неблагоприятные условия могут быть вызваны закупоркой канала ствола или патронника,
перегрузки пороха или из-за дефекта, неправильного или
неправильно загруженные и собранные компоненты картриджа. Даже
сильнейшее огнестрельное оружие может быть взорвано в результате чрезмерного
давление. Чрезвычайно опасно использовать картридж,
давление больше, чем у патронов, заряженных до
отраслевые стандарты.
БОЕПРИПАСЫ (ПАТРОНЫ) УВЕДОМЛЕНИЕМы специально отказываемся ответственность за любой ущерб, телесные повреждения или смерть, произошедшие в связи или в результате использования EAA Corp. ружья с неисправными, нестандартными, восстановленными, ручными или перезаряженные боеприпасы, использованные патроны или патроны, отличные от заводские патроны, под которые огнестрельное оружие предназначалось изначально под патрон.
ПРЕДУПРЕЖДЕНИЕ О СМАЗКЕ Стрельба из ружья с маслом, смазкой или любым другим
другой материал, даже частично закрывающий канал ствола, может привести к
повреждение ружья и телесные повреждения стрелка и тех,
рядом, поблизости. Не распыляйте и не наносите смазку непосредственно на боеприпасы. Если
на пороховой заряд патрона воздействует смазка, это
может еще не полностью воспламениться от энергии капсюля и/или пороха
может быть достаточно, чтобы протолкнуть снаряд или пыж в канал ствола, где
он может застрять. Выстрел следующего патрона в
закупоренный канал ствола повредит ружье и может стать причиной
ранение стрелка и окружающих. Правильно используйте смазочные материалы.
Вы несете ответственность за надлежащий уход и техническое обслуживание вашего
огнестрельное оружие и боеприпасы.
20
ОГРАНИЧЕННАЯ ГАРАНТИЯ НА ОДИН ГОД На это огнестрельное оружие EAA Corp.
первоначальному розничному покупателю в течение одного года с даты покупки
против дефектов материала и изготовления. Все детали и работа или
замена по нашему выбору покрываются. Деревянный приклад и деревянное цевье
не покрываются годовой гарантией. Гарантия на древесину
приклад и цевье 60 дней с момента покупки и покрывают только
производственные и материальные дефекты. Транспорт до и от нашего
средства ремонта, государственные пошлины, ущерб, причиненный несоблюдением
выполнять обычное техническое обслуживание, продажи за пределами США, повреждение
из-за использования высокой скорости, высокого давления, перезарядки или других
нестандартные боеприпасы или любой несанкционированный ремонт, модификация,
неправильное использование, злоупотребление или изменение продукта не подпадают под действие настоящего
Ограниченная гарантия. Любые подразумеваемые гарантии, включая подразумеваемые
гарантии товарного состояния и пригодности для конкретной цели,
ограничены одним годом с даты первоначальной розничной покупки.
Косвенные или случайные убытки и/или расходы, или любые другие
расходы не покрываются настоящей гарантией. Для получения гарантии
Performance отправьте свое огнестрельное оружие с подтверждением розничной покупки,
фрахт с предоплатой по адресу: European American Armory Corp. 3855 North U.S.
Шоссе 1 Какао, Флорида 32926
ВНИМАНИЕ: Незаконно перевозить огнестрельное оружие с боеприпасами в
огнестрельное оружие или в той же упаковке. Огнестрельное оружие и боеприпасы должны быть
поставляется отдельно. По вопросам доставки боеприпасов звоните по телефону
EAA Corp. по телефону (321) 639-4842 ПРЕДУПРЕЖДЕНИЕ ОБ ИЗМЕНЕНИЯХ ИЛИ
МОДИФИКАЦИИИзменение или модификация деталей и/или внутренних предохранительных устройств
опасно и приведет к аннулированию гарантии. Этот дробовик был
изготовлены для правильной работы с оригинальными деталями, как
разработан. Вы обязаны убедиться, что любые детали, которые вы покупаете, изготовлены
для этого огнестрельного оружия и установлены правильно и что ни
замены, ни оригиналы изменены или изменены. Ваш пистолет является
сложный прецизионный инструмент со многими частями, которые должны быть правильно связаны
к другим частям для правильной и безопасной работы. положить
пистолет вместе с неправильным или с неправильными или модифицированными частями может привести к
в поврежденном оружии, опасности и телесных повреждениях или смерти для вас и
другие из-за неисправности. Всегда есть квалифицированный оружейник
на вашем оружии или, по крайней мере, проверьте любую работу, не выполненную
оружейник.
ВАШЕ РУКОВОДСТВО ПО ЭКСПЛУАТАЦИИ Всегда храните это руководство вместе с вашим огнестрельным оружием.
Убедитесь, что вы понимаете все предупреждения, инструкции по эксплуатации
и процедуры безопасности. Когда вы одалживаете, дарите или продаете огнестрельное оружие,
уверен, что это руководство идет с ним. Вы можете получить копию этого руководства
от European American Armory Corp. по запросу.
21
ПРИ ЗАКАЗЕ ВСЕХ ЗАПЧАСТЕЙ ОПЛАЧИВАЕТСЯ 10 ДОЛЛАРОВ США ЗА ДОСТАВКУ И УМЕНИЕ ОБРАЩАТЬСЯ. Укажите модель, калибр, серийный номер и отделку. Цвет при заказе. Запчасти, информация и сервисное обслуживание: European American Armory Corp., 402 Richard Road, Rockledge, FL. 32955 Телефон: (321) 639-4842.
22
ПЕРЕЧЕНЬ ДЕТАЛЕЙ Рис. 1 ИЖ37 в разобранном виде
Список компонентов ИЖ371. ствол в сборе 2. выбрасыватель 3. винт
4. мушка 5. поворотное кольцо 6. штифт 7. шарнир в сборе 8. цевье
9. винт 10. втулка 11. корпус защелки 12. защелка передней рукоятки 13. штифт 14.
пружина 15. корпус 16. стопорный рычаг 17. винт 18. стопорный рычаг
ось 19. стопорная планка 20. возвратная пружина 21. верхний боек 22. нижний
ударник 23. пружина ударника 24. узел правого ударника 25.
левый ударник в сборе 26. штифт 27. шарнир главной пружины 28.
главная пружина 29. правое шептало 30. левое шептало 31. штифт 32.
пружина 33. толкатель правого болта 34. толкатель левого болта 35.
кнопка предохранителя 36. основание предохранителя 37. предохранитель 38. штифт 39.
пружина 40. пружина 41. зажим 42. штифт 43. ползунок 44. интерцептор 46.
пружина 47. винт 48. перемычка 49. опорная пластина 50. винт 51. блокировка
планка фиксатора 52. пружина 53. штифт 54. правый спусковой крючок 55.
левый спусковой крючок 56. штифт 57. спусковой крючок 58. курок 59. штифт 60.
спусковая скоба 61. шуруп 62. приклад 63. винт 64. шайба 66. приклад
пластина 67. Шуруп 68. Шуруп 69. поворотное основание 70. винт 71.
винт 72. Шайба
23
Рис. 2 — Детали выталкивателя ИЖ37
Рис. 2- ПЕРЕЧЕНЬ КОМПЛЕКТУЮЩИХ1. ствол в сборе 2. правый
выталкиватель 3. левый выбрасыватель 4. пружина выбрасывателя 5. плунжер 6. шарнир
узел 7. цевье 8. правое шептало выбрасывателя 9. выбрасыватель
левое шептало 10. болт шептала выбрасывателя 11. пружина 12. основание 13.
толкатель правого болта 14. толкатель левого болта 15. расцепление
рычаг 16. защелка рычага расцепления 17. пружина
24
Рис. 3 — Детали односпускового механизма ИЖ37
Рисунок 3- ПЕРЕЧЕНЬ КОМПОНЕНТОВ1. носитель 2. фиксирующая планка 3. правое шептало 4. левое шептало 5. рычаг инерционного расцепления 6. интерцептор 7. опорная плита 8. спусковой крючок 9. рычаг селектора 10. селектор пружина рычага 11. штифт рычага селектора 12. спусковой крючок 13. расцепитель пружина рычага 14. пружина спускового крючка 15. палец расцепляющего рычага 16. штифт спускового крючка 17. пружина спускового крючка 18. винт пружины рычага переключения передач 19. Пружина интерцептора
25
ДОПОЛНИТЕЛЬНЫЕ ДРОССЕЛЬНЫЕ ТРУБКИ ПРЕДУПРЕЖДЕНИЕ. Никогда не устанавливайте и не снимайте дроссель трубки во время загрузки огнестрельного оружия. Устанавливайте или удаляйте только чок трубки с незаряженным огнестрельным оружием, открытым затвором и спусковым крючком блокировка безопасности включена. Вверните символы/маркировку дросселей (это опция не все ружья ИЖ37 имеют эту функцию) Конец дульные насадки помечены косой чертой для идентификации чока сужение: / — Полное // — Улучшенное Модифицированное /// — Модифицированное //// — Улучшенный цилиндр Cl — цилиндр
ВНИМАНИЕ! УБЕДИТЕСЬ, ЧТО ВЫ ПОЛНОСТЬЮ ЗАТЯНУЛИ ДРОССЕЛЬНУЮ ТРУБКУ Стрельба
с незакрепленным дульным сужением может привести к повреждению ружья. Дополнительная информация о винтовых дульных сужениях: A) Затягивайте только
ослабьте дульные насадки с помощью ключа с плоским ключом, входящего в комплект ружья.
Примечание: ключ (и дополнительная дульная трубка или трубки, если они поставляются) находятся под
картон перед ствольной коробкой дробовика в коробке. Б) Никогда
стрелять из дробовика с частично установленной дульной насадкой. В) ИЖ37
ружья доступны с фиксированным дульным сужением или винтовым дульным сужением.
Доступны два типа винтовых дросселей: дроссель со свинцовой дробью
трубки и дульные насадки со стальной дробью. При установке стальной дроссель
трубы выступают примерно на 3/4 из дульного среза ствола
и сопрягаемая кромка будет на одном уровне с концом ствола. Когда
установлены, дульные насадки со свинцовой дробью устанавливаются заподлицо или чуть ниже
дульный конец ствола. Если у вас есть свинцовые дульные сужения и
намереваетесь стрелять патронами со стальной дробью или пулями, то вы должны использовать
только трубки IC или Cylinder. ЗАПРЕЩАЕТСЯ стрелять стальной дробью или пулями в
Модифицированные, улучшенные модифицированные или полностью свинцовые дульные насадки. если ты
иметь дульные сужения со стальной дробью, вы можете стрелять патронами со стальной дробью
через любую из этих дроссельных трубок. НЕ стреляйте пулями в Modified,
Улучшенные модифицированные или полностью свинцовые дульные насадки или дульные сужения со стальной дробью
трубы.
26
D) Правильно установленные дульные насадки: дульные насадки со свинцовой дробью должны
устанавливаться заподлицо или чуть ниже дульного среза ствола. Стали
дульные насадки будут выступать из ствола примерно на 3/4 и
сопрягаемая кромка будет на одном уровне с концом ствола. Е) Перед использованием
Убедитесь, что дульные насадки установлены правильно. Ф)
Держите дульные насадки смазанными и установленными. Никогда не стреляйте из пистолета
без установленной дульной насадки. Аккуратно обращайтесь с дульными насадками, т.к.
передняя кромка очень тонкая. Примечания по материалу дроби и чокам:
1) Снаряды с висмутовой или вольфрамовой матрицей подлежат
те же ограничения ствола и дульной насадки, что и у свинцовой дроби. Оболочки с использованием
Выстрелы из TungstenIron или Hevi-Shot относятся к одному и тому же стволу и
ограничения дульной насадки в виде стальной дроби. 2) На ружьях с фиксированными чоками,
снаряды, заряженные сталью, вольфрамовым железом или HeviShot, должны стрелять
только в стволах IC или Cylinder. ЗАПРЕЩАЕТСЯ сжигать сталь, вольфрам-железо или
Снаряды Hevi-Shot в несъемных стволах с модифицированным, импровизированным модифицированным
или Полные чоки. Свинец, висмут и вольфрамовая матрица могут быть
стрелял из всех стволов с фиксированным дульным сужением. 3) На фиксированных дульных ружьях пули могут
стрелять только из стволов IC и Cylinder. НЕ стреляйте пулями в
фиксированные дульные сужения с модификацией, улучшенной модификацией или полной
душит. Если вы хотите купить дополнительные дульные насадки (скит, трап,
дополнительный полный и т. д.) звоните 1-321-639-4842
27
!
ПРЕДУПРЕЖДЕНИЕ
Детей привлекает огнестрельное оружие, которое может
привести к тяжелым травмам или смерти. Всегда запрещайте доступ детям
держать оружие запертым и разряженным, когда оно не используется. Если вы держите
заряженное огнестрельное оружие, которое ребенок получает и использует ненадлежащим образом, вы
может быть оштрафован или отправлен в тюрьму.
!
ADVERTENCIA
A los ninos los atroen las armaside de fuego y los pueden hacer функциональный. Ellos pueden causarses lesiones могилы и ла muerte. Evite Que los ninos tengon acceso a los armas de fuego guardandolas siempre con llave y descargadas cuando no las este utilizando. Си usted tiene un arma de fuego cargado en un lugar en que un nono tiene acceso a ella y la usa indebidamente. Le pueden dar una multa enviarlo a la carcel.
Импортер: European American Armory Corp. Оружейная палата 3855402 RichardHighway 1 North US Road Cocoa, FL 32926 Рокледж, Флорида. 32955 Производитель: Ижевский Механический Завод 8, ул. Промышленная, 426063 Ижевск, Россия
Четыре коробчатых ружья с вертикальным расположением стволов -A) Байкал Модель ИЖ 27ЕМ-1С-М Вертикальное ружье с коробкой B) Khan A
A) Стандартная маркировка и характеристики. Под патроны 3 дюйма. Длина тяги 14 1/4 дюйма. В комплекте оригинальная коробка с номером ружья, 3 дульных насадки, дульный инструмент и документы.
BBL: вентиляционное ребро 27 3/4 дюйма
Ложа: орех в клетку
Толщина: 12
Отделка: синяя
Рукоятки:
Серийный номер: 9840248B
Состояние: новое — все оригинальные детали; 99% или более оригинальная отделка.
B) Стандартная маркировка и характеристики. Рассчитан на 3-дюймовые снаряды. Длина тяги 14 1/4″. В комплекте оригинальная коробка с номером пистолета, 5 дульных насадок, дульный инструмент и документы.
BBL: вентиляционная планка 28 дюймов
Ложа: орех с насечкой
Калибр: 12
Отделка: синяя
Захваты:
Серийный номер: TR12100433
Состояние: новое — древесина хуже по проценту отделки Дан
C) Представлен в 2005 году, со стандартной маркировкой и функциями. Под патроны 2 1/2 дюйма и 3 дюйма. Длина тяги 14 дюймов. В комплекте оригинальная коробка с номером пистолета и документами.
BBL: вентиляционная планка 26 дюймов
Ложа: орех
Калибр: 410
Отделка: синяя/сатиновая монета
Рукоятки:
Серийный номер: TR410003293
Состояние: Отличное 90%-94%
D) Представлен в 2005 г., со стандартной маркировкой и характеристиками. Предназначен для патронников 2 3/4″ и 3″. Длина тяги 14″. В комплекте оригинальная коробка с номером пистолета, 5 дульных насадок, дульный инструмент и документы.
BBL: 26-дюймовое вентиляционное ребро
Ложа: клетчатый орех
Толщина: 20
Отделка: синяя/атласная монета
Рукоятки:
Серийный номер: TR20007783
Состояние: новое — все оригинальные детали; 99% или более оригинальная отделка.
Место аукциона:
4507 49th Avenue, Молин, Иллинойс, 62165, США
Детали предварительного просмотра:
Пятница, 25 июня, с 10:00 до 18:00.
Суббота, 26 июня с 8:00 до 10:00.
(полный предварительный просмотр предметов, продаваемых в воскресенье)
Воскресенье, 27 июня с 8:00 до 10:00.
(для товаров, продаваемых в воскресенье)
Премии покупателя:
Дополнительные сборы:
| Нет данных. |
Детали доставки:
После аукциона с вами свяжутся для согласования оплаты и доставки. Стоимость доставки будет добавлена к вашему счету.
Детали платежа:
Информация об оплате: Аукционные продажи осуществляются строго за наличные, банковский чек или чеки (с предварительного одобрения руководства), карты MasterCard, Visa или Discover. ПРИМЕЧАНИЕ ДЛЯ ВСЕХ ПОКУПАТЕЛЕЙ. Способ оплаты: валюта США или заверенный чек, выписанный в американском банке. Все продажи являются окончательными при падении молота аукциониста. Премия покупателя: все цены предложения «цены молотка» (означает цену, по которой лот сбивается покупателю) облагаются премией покупателя, уплачиваемой покупателем в размере 19. 5% от цены предложения. При оплате наличными, утвержденным чеком или банковским переводом предоставляется скидка 2,5%. Премия добавляется к вашей успешной ставке, и вместе они равняются общей цене покупки. При объявлении «ПРОДАНО» предложенный лот приобретет тот, кто предложит самую высокую цену. На эту покупку распространяются все положения и условия, изложенные в настоящем документе. Покупатель берет на себя полную ответственность за это и, если потребуется, подпишет подтверждение покупки. Покупатель также соглашается уплатить покупную цену, премию покупателя и любые применимые налоги до того, как какой-либо лот может быть передан. Все товары, приобретенные победителями торгов, должны быть полностью оплачены в день продажи. Все товары, приобретенные участниками заочных торгов, должны быть полностью оплачены сразу после уведомления.
Принятые способы оплаты:
- VISA
- Мастер-карта
- Откройте для себя
- Денежный перевод/банковская тратта
- Чек/Чек
- См. реквизиты платежа
- Банковский перевод
1. Следующие условия продажи представляют собой полные условия, на которых предметы, перечисленные в нашем каталоге и дополнительных листах, будут предлагаться для продажи или продаваться аукционной компанией Rock Island и любым клиентом, для которого мы выступаем в качестве агента. Любые уведомления, размещенные или устные во время продажи, также являются частью нашего условия договора купли-продажи.
2. Все цены предложения «цены молотка» (означает цену, по которой лот сбивается покупателю) облагаются премией покупателя, уплачиваемой покупателем в размере 19,5% от цены предложения (19,5% при использовании Visa, MasterCard или Discover за наличные или утвержденный чек, при этом надбавка покупателя составит 17%.) Премия добавляется к вашей успешной ставке, и обе суммы вместе составляют общую цену покупки. Некоторые товары освобождены от налога с продаж штата Иллинойс, а другие могут облагаться налогом с продаж штата Иллинойс или местным налогом с продаж. Налог рассчитывается от общей стоимости покупки. Чтобы установить безналоговую продажу, необходимо предоставить копию действительного разрешения на реселлеров или другой документ, подтверждающий освобождение от налога с продаж. Любой покупатель, претендующий на освобождение, но не способный предоставить доказательства, должен будет заплатить нам налог и потребовать возмещения от штата Иллинойс. Как правило, все товары, отправляемые за пределы штата, не облагаются налогом. Грузоотправитель уполномочил нас удерживать в качестве части нашего вознаграждения надбавку, подлежащую уплате покупателем.
3. Продажи на аукционе строго за наличные деньги, кассовый чек или чеки (с предварительного согласования с руководством), карты MasterCard, Visa или Discover. ПРИМЕЧАНИЕ ДЛЯ ВСЕХ ПОКУПАТЕЛЕЙ. Способ оплаты: валюта США или заверенный чек, выписанный в американском банке. Все продажи являются окончательными при падении молота аукциониста.
4. При объявлении «ПРОДАНО» предложенный лот приобретет участник, предложивший самую высокую цену. На эту покупку распространяются все положения и условия, изложенные в настоящем документе. Покупатель берет на себя полную ответственность за это и, если потребуется, подпишет подтверждение покупки. Покупатель также соглашается уплатить покупную цену, премию покупателя и любые применимые налоги до того, как какой-либо лот может быть передан. Все товары, приобретенные победителями торгов, должны быть полностью оплачены в день продажи. Все товары, приобретенные участниками заочных торгов, должны быть полностью оплачены сразу после уведомления. Все имущество должно быть вывезено из наших помещений за счет покупателя не позднее 17:00. через пять дней после даты продажи, если не оговорено иное. Если имущество не вывезено, оно может быть передано покупателю на общественный склад на риск и за счет покупателя. Каждый покупатель на этом аукционе предоставляет нам залог в приобретенном имуществе. Мы можем сохранить в качестве залога по обязательствам покупателя перед нами любое имущество покупателя и все денежные средства, находящиеся или полученные нами. Мы можем использовать в отношении таких обязательств денежные средства, удерживаемые или полученные нами за счет или причитающиеся покупателю. Мы сохраняем за собой все права обеспеченной стороны в соответствии с Коммерческим кодексом штата Иллинойс. Всякий раз, когда покупатель оплачивает только часть общей цены за один или несколько приобретенных лотов, мы можем применить такие платежи по нашему собственному усмотрению к выбранному нами лоту или лотам. Платеж не будет считаться произведенным в полном объеме до тех пор, пока мы не получим все суммы, причитающиеся наличными, на хорошие средства (по нашему определению). Если условия продажи не соблюдаются, в дополнение к средствам правовой защиты, доступным нам и грузоотправителю по закону, мы возлагаем на покупателя ответственность за уплату покупной цены, премии покупателя и применимого налога с продаж, мы по нашему выбору можем: (1) отменить продажу, сохранить в качестве неустойки все платежи, сделанные покупателем, или (2) перепродать имущество либо публично, либо в частном порядке, и в таком случае покупатель несет ответственность за оплату любой недостачи плюс все затраты и расходы, связанные с обеими продажами. и наша комиссия по нашим стандартным ставкам. Покупатель также несет ответственность за все другие расходы, причитающиеся по настоящему Соглашению, а также гонорары адвокатов, случайные убытки и т. д.
5. Мы оставляем за собой право изъять любое имущество до продажи.
6. Мы оставляем за собой право отклонить предложение любого участника торгов. Покупателем станет тот, кто предложит самую высокую цену, признанную аукционистом. Аукционист имеет единоличное и окончательное усмотрение в отношении разрешения любого спора, включая повторное предложение и перепродажу любого оспариваемого предмета. Любой спор после продажи нашей записи будет считаться окончательным во всех отношениях.
7. Если пожар, кража или любая другая причина помешали нам передать какое-либо имущество покупателю, наша ответственность ограничивается суммой, фактически уплаченной покупателем, и ни в коем случае не включает любые случайные или косвенные убытки. .
8. Все имущество, выставленное на продажу, находится в том виде, в каком оно есть. Аукцион Rock Island не гарантирует и не дает гарантий ни на один проданный лот. Описания в каталоге являются мнением. Они написаны в качестве помощи для потенциальных участников торгов. Мы признаем, что в написанном могут быть ошибки. Суть в том, что если вы сделали ставку на него и выиграли, вы владеете им. Мы настоятельно рекомендуем вам лично просмотреть любой предмет, на который вы делаете ставку, или обратиться к признанному эксперту для осмотра предмета. Аукционная компания Rock Island не несет никаких обязательств, в том числе не принимает возмещений или возвратов. Предметы, предлагаемые для продажи, как описано в каталоге или любом чеке, рекламе, дополнительном листе или где-либо еще в отношении авторства, периода, культуры, источника, происхождения, размера, качества, редкости, происхождения, важности, выставки, физического состояния. заявления о мнении, а не заверения или гарантии. Ни один сотрудник аукционной компании Rock Island не уполномочен делать от нашего имени или от имени грузоотправителя какие-либо заверения или гарантии, устные или письменные, в отношении любого имущества.
9. Если упаковка и транспортировка купленных лотов осуществляется аукционной компанией Rock Island, это делается на риск покупателя. Все предметы будут взиматься за страхование; эта страховка обязательна. Мы не несем ответственности за какие-либо упущения при упаковке или доставке. Приобретенные партии, обрабатываемые сторонними перевозчиками или упаковщиками, в том числе теми, кого мы могли порекомендовать, могут иметь свою собственную страховку, и любые претензии в отношении убытков или повреждений должны быть адресованы непосредственно им.
10. Настоящие условия продажи, покупатели и наши соответствующие права и обязанности по ним регулируются законодательством штата Иллинойс. Принимая участие в торгах на нашем аукционе, будь то лично или через агента, запечатанные ставки, ставки по телефону или другими способами, покупатель или участник торгов соглашается с тем, что контракт, созданный в соответствии с настоящими условиями продажи, заключен и выполняется в округе Рок-Айленд, штат Иллинойс.
11. Условия продажи связывают правопреемников и правопреемников всех участников торгов и покупателей и действуют в интересах наших правопреемников и правопреемников. Никакие отказы, поправки или модификации условий настоящего Соглашения (кроме размещенных уведомлений или устных объявлений во время продажи) не обязывают нас, если это специально не указано в письменной форме, подписанной нами. Если какая-либо часть настоящих Условий продажи по какой-либо причине недействительна или не имеет исковой силы, остальные остаются в силе и подлежат исполнению.
12. На некоторые лоты данного аукциона может распространяться резерв (конфиденциальная минимальная цена, ниже которой лот не будет продан). В соответствии с политикой сумма резерва не будет превышать нижнюю оценку и во многих случаях может быть фактически меньше нижней оценки. Если лот предлагается с резервом, мы можем реализовать этот резерв на торгах от имени грузоотправителя. Эти торги не будут считаться открытыми торгами. Если не будет получена начальная ставка, мы пропустим этот пункт. Однако, как только торги будут открыты, мы подадим заявку от имени грузоотправителя, чтобы достичь резервной цены. Если у нас есть интерес к предлагаемому лоту и поступлениям от него, кроме нашей комиссии, мы можем подать заявку, чтобы защитить такой интерес. Предметы, которые не зарезервированы, продаются по усмотрению аукциониста.
13. Грузоотправителям не разрешается делать ставки на свои собственные товары или привлекать агентов, делающих ставки от их имени. Если Аукционист узнает о таких торгах или ему сообщают об этом, Аукционист оставляет за собой право отозвать любые или все предметы, переданные нарушившим отправителем, без каких-либо исключений.
14. В качестве услуги для всех, кто желает сделать ставки до начала продажи, мы можем принимать ставки по своему усмотрению по телефону или запечатывать ставки, используя формы, которые мы предоставляем. Торги по телефону в региональных продажах разрешены только в особых случаях. Телефонные торги в региональных продажах ограничены. Напоминаем всем заочным участникам этой региональной распродажи, что все распродажи являются окончательными. Запечатанная или телефонная ставка должна быть получена и одобрена кредитом до даты продажи. Ответственность за установление кредита перед торгами лежит на участниках торгов, в противном случае мы примем 15% депозита от суммы всех предложений. (Депозиты будут возвращены в течение десяти (10) дней после продажи, в случае неудачи). Форма закрытой заявки прилагается в конце каталога. Тендерные предложения также можно отправить по факсу (309)797-1655. Мы не несем ответственности за невыполнение телефонных или запечатанных заявок по какой-либо причине.
15. Все огнестрельное оружие выпуска 1898 года должно быть зарегистрировано в соответствии с Федеральным законодательством и законодательством штата Иллинойс. Покупатели огнестрельного оружия после 1898 года должны заполнить формы государственной и федеральной регистрации в аукционной компании Rock Island, 4507 49th Avenue, Moline, Illinois 61265, если не указано иное. 3-дневный период ожидания является обязательным для современного пистолета, 24-часовой период ожидания для современного длинноствольного оружия. Дилеры и покупатели за пределами штата должны иметь при себе в день продажи подписанные копии своей федеральной лицензии на огнестрельное оружие, чтобы принять поставку современного оружия в тот же день. Покупатели, выступающие в качестве агентов дилеров FFL, должны иметь представительское письмо, а также оригинал подписанной копии Федеральной лицензии дилера на огнестрельное оружие (FFL).
16. Аукционная компания Rock Island не дает никаких гарантий или заявлений, и ни один сотрудник или консультант аукционной компании Rock Island не имеет полномочий делать иное в отношении эксплуатации или условий обжига, пригодности к использованию, безопасности хранения или надежности любого огнестрельного оружия, боеприпасов или частей. Использование любого огнестрельного оружия или боеприпасов, приобретенных на аукционе Rock Island Auction Company, осуществляется исключительно на риск пользователя. Аукционная компания Rock Island выставляет лоты на продажу только как «коллекционные». Мы настоятельно рекомендуем, чтобы все оружие, боеприпасы и т. д., приобретенные на аукционе, были проверены компетентным оружейником. Аукционная компания Rock Island прямо отказывается от какой бы то ни было ответственности за несчастный случай, травмы или ущерб, нанесенный любому лицу в результате хранения или последующего использования любого такого лота.
17. Все расходы по доставке, отгрузке, страхованию и хранению должны быть оплачены победителем торгов до поставки огнестрельного оружия.
18. Мы организуем упаковку и отправку в кратчайшие сроки. Мы постараемся отправить товар как можно скорее, однако из-за большого количества заочных участников торгов задержка доставки в течение семи дней после получения нами полной оплаты является типичной. В случае упаковки в ящики или любой другой необычной упаковки покупатель будет нести нашу стоимость от сторонних агентов. Покупатель оплачивает все расходы по доставке и страховке.
19. Принятие карты участника торгов вместе с признанной ставкой означает принятие вышеупомянутых условий продажи.
20. Ограничение ответственности. все имущество продается «как есть». Аукционная компания Rock Island, ни грузоотправитель не делают никаких заявлений или гарантий, явных или подразумеваемых, в отношении названия, пригодности для продажи, пригодности или состояния имущества или в отношении правильности описания, подлинности, атрибуции, происхождения или периода собственности или от того, приобретает ли покупатель какие-либо авторские права или другие права интеллектуальной собственности на проданные лоты, или от того, подчиняется ли произведение искусства неимущественным правам художника или другим остаточным правам художника. Все огнестрельное оружие и боеприпасы продаются только коллекционными лотами. Аукционная компания Рок-Айленда категорически отрицает пригодность для стрельбы или иного разряда оружия или боеприпасов. следует проконсультироваться с компетентным оружейником, прежде чем пытаться использовать или стрелять из любого огнестрельного оружия или боеприпасов. покупатель прямо признает и соглашается с тем, что ни при каких обстоятельствах аукционная компания Rock Island ИЛИ ГРУЗОПОДЪЕМНИКИ не несут ответственности за любые убытки, включая, помимо прочего, любые компенсационные, случайные или косвенные убытки.
21. Телефон. Ставки по телефону будут ограничены. Мы принимаем только небольшое количество предложений по телефону. Все торги по телефону будут основываться на истории прошлых покупок клиентов в аукционной компании Rock Island. Первоначально региональные продажи должны были быть ограничены, а торги по телефону не разрешались. Теперь мы признаем, что могут возникнуть особые обстоятельства. Мы постараемся учесть наших бывших клиентов в особых обстоятельствах.
Полногеномное секвенирование изолята Borrelia miyamotoi Иж-4: эталон сложного бактериального генома | Геномика BMC
- Исследовательская статья
- Открытый доступ
- Опубликовано:
- Кулешов Константин Владимирович Orcid: orcid.org/0000-0002-5238-7900 1,2 ,
- Габриэль Маргос 3 ,
- Volker Filmle 3 ,
- Joris Koetselld 4 ,
- Joris Koetselld 4 ,
- A. , ,
- A. , ,
- A. 4 ,
- A. 4 ,
- A. 4 ,
- .2315 5 ,
- Mikhail L. Markelov 5 ,
- Nadezhda M. Kolyasnikova 1,6 ,
- Denis S. Sarksyan 4,7 ,
- NINA P. KIRINASHINASHINASHINASHINASHINASHINASHINA. Шипулин Герман А. 8 ,
- Ховиус Йоппе В. 4 и
- …
- Платонов Александр Евгеньевич 1
Геномика BMC том 21 , Номер статьи: 16 (2020) Процитировать эту статью
2903 Доступы
14 цитирований
21 Альтметрический
Сведения о показателях
Abstract
История вопроса
Род Borrelia включает спирохетальные бактерии, поддерживаемые в естественных циклах передачи клещами-переносчиками и позвоночными хозяевами-резервуарами. Основные группы представлены видовым комплексом, включающим возбудителей Лайм-боррелиоза и возвратного тифа группу Borrelia . Borrelia miyamotoi принадлежит к группе спирохет возвратного тифа и образует отдельные популяции в Северной Америке, Азии и Европе. Как и все Borrelia видов B. miyamotoi обладают необычным и сложным геномом, состоящим из линейной хромосомы и ряда линейных и кольцевых плазмид. Этот вид считается новым патогеном человека, и в Северном полушарии описывается все большее число случаев заболевания людей. Целью этого исследования было создание высококачественного эталонного генома, который облегчит будущие исследования генетических различий между различными популяциями и пластичности генома 9 видов.2394 Б. Миямотои .
Результаты
Мы использовали несколько доступных методов секвенирования, в том числе одномолекулярную технологию Pacific Bioscience в режиме реального времени (SMRT) и технологию Oxford Nanopore (ONT), дополненную высокоточными последовательностями Illumina, чтобы изучить пригодность для сборки всего генома российского Изолят B. miyamotoi , Иж-4. Плазмиды типировали в соответствии с их потенциальными генами разделения плазмид (PF32, 49, 50, 57/62). Сравнивая и объединяя результаты как длинного (SMRT и ONT), так и короткого (Illumina) методов, мы определили, что геном изолята Иж-4 состоит из одной линейной хромосомы, 12 линейных и двух кольцевых плазмид. В то время как большинство плазмид имели соответствующие контиги в азиатских B. miyamotoi изолят FR64b, было только четыре, которые соответствовали плазмидам североамериканского изолята CT13–2396, что указывает на различия между популяциями B. miyamotoi . Несколько плазмид, например Было обнаружено, что lp41, lp29, lp23 и lp24 несут вариабельные основные белки. Среди них были вариабельные большие белки (Vlp) подтипов Vlp-α, Vlp-γ, Vlp-δ, а также Vlp-β. Филогенетический анализ распространенных типов плазмид показал уникальность российских/азиатских изолятов B. miyamotoi по сравнению с другими изолятами.
Выводы
Здесь мы описываем геном российского клинического изолята B. miyamotoi , обеспечивая прочную основу для будущей сравнительной геномики изолятов B. miyamotoi . Это станет большим стимулом для дальнейших фундаментальных, молекулярных и эпидемиологических исследований этого нового переносимого клещами патогена.
Исходная информация
Borrelia miyamotoi была впервые обнаружена в Ixodes persulcatus в Японии и описана в 1995 [1]. Впоследствии было обнаружено, что он встречается симпатрически с B. burgdorferi sensu lato у нескольких видов Ixodes , которые также являются переносчиками спирохет болезни Лайма. К ним относятся Ixodes persulcatus в Евразии [2,3,4,5,6,7], I. scapularis [8,9,10,11] и I. pacificus [12,13,14,15]. ] в Северной Америке и I. ricinus в Европе [16,17,18,19,20]. Установлено, что распространенность B. miyamotoi среди клещей обычно ниже, чем у B. burgdorferi с.л. хотя в некоторых регионах сообщалось о распространенности ~ 15% [3, 7, 10, 16, 17, 21, 22]. Предполагается, что грызуны являются резервуарными хозяевами B. miyamotoi [23, 24], но также известно, что происходит трансовариальная передача [25, 26], что может способствовать сохранению этого Borrelia в природе.
Несмотря на совместное появление с B. burgdorferi s.l. у твердотелых иксодовых клещей , генетический и филогенетический анализы показали, что B. miyamotoi относится к кладе спирохет возвратного тифа (РФ) [1, 2, 16, 23, 27], переносчиками которых обычно являются мягкие клещи ( Argasidae ) или вши. Подобно другим видам возвратного тифа, B. miyamotoi обладает генами, кодирующими вариабельные большие белки и вариабельные малые белки (Vlp и Vsp соответственно) [11, 28, 29]. Vlp и Vsp экспрессируются во время позвоночной фазы жизненного цикла спирохет возвратного тифа. Эти белки принадлежат к системе антигенной изменчивости спирохет, которая позволяет избежать приобретенного иммунного ответа хозяина. Это может пролонгировать присутствие спирохет в кровотоке инфицированного животного, увеличивая тем самым возможность передачи переносчику [30, 31]. Генетические исследования образцов, собранных в полевых условиях, показали, что существует небольшая генетическая изменчивость изолята B. miyamotoi в популяции одного вида клещей, тогда как изолята B. miyamotoi от разных видов клещей оказались генетически гетерогенными [3, 22]. Так, было высказано предположение, что вид B. miyamotoi состоит из азиатского, европейского, североамериканского — западного и восточного побережья — экотипов/генотипов [2, 8, 16, 32, 33].
Первые случаи заболевания человека, вызванного B. miyamotoi , зарегистрированы в 2011 г. в России [3]. В этом исследовании 46 случаев из 9Было описано 2394 болезни B. miyamotoi (BMD) с клиническими проявлениями, которые включали лихорадку и гриппоподобное заболевание, с миалгией и артралгией среди других симптомов. С тех пор в России было выявлено несколько сотен случаев МПК [34, 35]. Случаи МДБ были зарегистрированы также в Европе и США, но не с такой частотой [2, 36,37,38,39]. Случаи, о которых сообщалось в Западной Европе, часто касались лиц с ослабленным иммунитетом, но в последнее время также и иммунокомпетентных лиц [40, 41]. Широкое географическое распространение этого нового патогена человека, который может использовать множество различных переносчиков и хозяев, а также различные клинические проявления МПК, варьирующие по клинической значимости от бессимптомной инфекции до тяжелых последствий, таких как менингоэнцефалит, подразумевают необходимость понимания генетической основы. этого разнообразия.
Однако по сравнению с геномами других бактерий геномы Borrelia необычайно сложны и состоят из линейной хромосомы и ряда линейных и кольцевых плазмид. Содержание и структура плазмид различаются не только среди видов, но и внутри видов. Таким образом, сборка полного генома B. miyamotoi является сложной задачей.
На данный момент геном одного изолята B. miyamotoi FR64b азиатского подтипа и четырех американских изолятов (CT13–2396, CA17–2241, LB2001, CT14D4) были секвенированы [11, 14, 33, 42]. Однако метод длительного секвенирования использовался только для характеристики CT13–2396. Поэтому количество и содержание плазмид не описано должным образом для остальных четырех штаммов [43].
В текущем исследовании мы секвенировали геном одного российского пациента, изолированного B. miyamotoi . Целью нашего исследования было создание генома высокого качества для B. miyamotoi , чтобы обеспечить основу для дальнейших исследований генетического разнообразия и пластичности генома 9.2394 Б. Миямотои . С этой целью мы оценили несколько методов секвенирования и биоинформатики, а также несколько методов идентификации и классификации плазмид. Мы сравнили и объединили различные методы длительного считывания (технология одиночных молекул в реальном времени Pacific Biosciences (SMRT) и технология Oxford Nanopore (ONT)) и дополнили сборки точными последовательностями короткого считывания Illumina. Полученный эталонный геном поможет упростить и улучшить будущий геномный анализ B. miyamotoi 9.2367, в частности, для изучения специфических геномных особенностей азиатских изолятов B. miyamotoi , а также для выявления и изучения факторов вирулентности и патогенности.
Результаты
Анализ PFGE
B. miyamotoi Штамм Иж-4Гель-электрофорез в импульсном поле (PFGE) выявил хромосому длиной ~ 900 кб и девять нехромосомных фрагментов (потенциальные плазмиды) (рис. . 1). Первые три нехромосомных фрагмента размером от 72 кб до 64 кб были сходны у всех русских изолята B. miyamotoi [44] (данные не показаны). Остальные полосы указывали на присутствие дополнительных шести плазмид размером от прибл. от 40 кб до 13 кб. Вероятно, это заниженная оценка, поскольку хорошо известно, что плазмиды с одинаковыми размерами или кольцевые плазмиды (которые могут иметь другие модели миграции, чем линейные плазмиды) не могут быть идентифицированы с помощью PFGE.
Рис. 1 ПФГЭ-спектр хромосомной и плазмидной ДНК изолята B. miyamotoi Иж-4 в трех независимых повторах. N1-N9указывают фрагменты PFGE, которые были подвергнуты гелевой экстракции и секвенированию с помощью платформы Illumina. Название плазмид соответствующей длины указано на правом участке геля. Он был основан на сравнении собранных контигов из каждого из фрагментов PFGE с окончательной сборкой. Следует отметить, что плазмида lp6 не разделялась в PFGE, не было видно отчетливой полосы такого размера. Это могло быть связано с недостаточными условиями PFGE, так как последовательности lp6 были идентифицированы во фрагменте размером 13 т.п.н. вместе с плазмидой lp13 путем прямого секвенирования
Изображение в натуральную величину
Штамм B. miyamotoi , секвенирование и сборка генома Для получения высококачественного эталонного генома для сравнительной геномики B. miyamotoi геном изолята Иж-4 был выбран случайным образом из доступные российские клинические изоляты [44] ( Дополнительный файл 1: Таблица S1 ) и секвенированы с использованием различных платформ секвенирования, включая Illumina MiSeq и HiSeq, ONT MinION и Pacific Biosciences SMRT. Сборки длинных ридов были исправлены с помощью длинных ридов (например, PacBio с PacBio; ONT с ONT) и впоследствии с использованием высокоточных чтений последовательностей Illumina с помощью конвейера Pilon [45].
Используя платформу MinION, мы получили 129 992 необработанных чтения средней длины 6,6 КБ. После исправления и обрезки в конвейере Canu v1.7 количество длинных чтений уменьшается до 31 584 при средней длине 7,3 кб. Сборка показала 16 контигов длиной от 900 kb до 10 kb. Ручная валидация показала, что два из них — tig00009030 и tig00000013 — характеризовались специфическим паттерном покрытия прочтений ONT в двух пиках, указывающих на слияние двух отдельных плазмид. Кроме того, два контига имели размер 46 КБ и 50 КБ, что не соответствовало анализу PFGE (дополнительный файл 2: рисунки S1–S3). Поэтому эти контиги были разделены на два контига и обработаны как отдельные плазмиды. Кроме того, три из полученных 18 контигов характеризовались низким охватом длинным чтением (2-3x) и имели высокий уровень сходства (≥ 9). 5%) на другие контиги и поэтому были исключены из дальнейшего анализа. Наконец, два из 15 оставшихся контигов были автоматически циркулярированы с длиной 30 kb и 29 kb. Подводя итог, с помощью этого метода в итоге мы получили 15 контигов, соответствующих одной основной хромосоме, и 14 потенциальных плазмид с охватом обрезанными ридами в диапазоне от 300x до 20x (таблица 1).
Полноразмерная таблица
Используя платформу PacBio, мы получили 312 224 необработанных чтения со средней длиной 4 КБ. Используя 2635 исправленных прочтений со средней длиной 8,8 т.п.н., было собрано 20 контигов с длиной контигов от 6 до 906 т.п.о. Три контига с низким покрытием, с последовательностями, присутствующими в других частях генома, были сочтены артефактами сборки и были удалены. Два контига были вручную округлены на основе перекрывающихся концов.
Были отмечены несоответствия между сборками ONT и PacBio, а также различия в гипотетических длинах плазмид в PFGE. PacBio unitig#3 имел размер 68 kb и не был идентифицирован в PFGE. Это было похоже на три отдельных контига ONT (41 КБ, 27 КБ и 22 КБ) (дополнительный файл 2: рисунок S4). Были идентифицированы три блока PacBio, соответствующие контигу ONT размером 70 КБ, поэтому контиг ONT был ошибочно разделен на три отдельных контига PacBio (дополнительный файл 2: рисунок S5). Более того, две из этих единиц PacBio # 20 (~ 38 kb) и № 22 (~ 38 kb) не наблюдались в PFGE. Контиг ONT размером 64 КБ был частично представлен в блоке № 10, который имел размер 43 КБ (дополнительный файл 2: рисунок S6) и также не был найден в PFGE. Эти неправильные сборки последовательностей PacBio могли быть вызваны малым количеством ДНК, представленным для секвенирования (1,2 мкг), что было меньше, чем требовалось службой секвенирования (5–10 мкг), и не позволяло выбрать размер BluePippin. Тем не менее, остальные контиги были одинаковыми между сборками PacBio и ONT. Контиги ONT, которые были разделены на основе анализа покрытия, были подтверждены юнитигами PacBio как отдельные последовательности. В целом, извлеченные консенсусные последовательности из сборок PacBio и ONT (скорректированные с помощью высокоточных считываний Illumina) привели к полному геному, состоящему из хромосомы ~ 900 kb, и 14 предполагаемых плазмидных контигов, из которых два были круглыми и 12 линейными, длиной от 6 до 73 kb.
Контиги вышеописанной окончательной сборки также сравнивали с контигами, полученными прямым секвенированием фрагментов ДНК, извлеченных из агарозного геля после разделения с помощью PFGE. Эти контиги были сопоставлены с помощью Mummer и визуализированы с помощью Circos. Для разных полос было получено несколько контигов, но только подмножество в каждой полосе представляло рассматриваемую плазмиду (см. рис. 1 и дополнительный файл 2: рисунки S7-S15). Например, для фрагмента PFGE N1 было собрано 85 контигов из коротких прочтений Illumina, но только один контиг длиной 72 707 bp полностью воспроизводил плазмиду lp72 в окончательной сборке. Хотя нам удалось идентифицировать большинство линейных плазмид путем прямого секвенирования фрагментов PFGE, среди собранных контигов не было обнаружено последовательностей, соответствующих кольцевым плазмидам (ср30-1 и ср30-2). Две плазмиды, а именно lp70 и lp64, были сильно фрагментированы. Наблюдалось множество небольших контигов с низким покрытием k-mer по сравнению с основными контигами, что, возможно, было результатом загрязнения образца в процессе выделения ДНК.
Окончательный состав генома представлен в таблице 1. Эта сборка была депонирована в GenBank, BioSample SAMN07572561.
Определение последовательностей теломер на левом и правом концах линейных репликонов
Геном изолята Иж-4 Borrelia miyamotoi содержит 13 линейных репликонов. Поскольку сообщалось о палиндромных последовательностях на концах линейных плазмид у других видов Borrelia [46], мы искали, были ли линейные репликоны фланкированы палиндромными последовательностями, которые напоминают короткие теломерные структуры, образующие ковалентно замкнутые шпильки. При анализе терминальных участков собранной хромосомы и линейных плазмид были выявлены терминальные последовательности нуклеотидов, которые представлены в таблице 2. Идентичные палиндромные последовательности обнаружены для lp70R и lp18–1 L, lp70L и lp13L, lp64L и lp41L, lp29.R/lp24L/lp23R, lp29L и lp27L, lp24R и lp18–2 L. Последовательность lp6L, хотя и палиндромная, могла быть не идентифицирована должным образом, поскольку не было «сигнатурной» последовательности.
Полная таблица miyamotoi , мы можем только предположить, что есть доказательства наличия «бокса 3» с консенсусным мотивом «WTWGTATA», начинающимся с позиции 14, как описано ранее для болезни Лайма 9.2394 Боррелия [46,47,48]. Последовательность, обозначенная как «Вставка 3», соответствует ранее аннотированной консервативной области (Вставка 3), которая, как предполагалось, непосредственно участвует во взаимодействии с теломерной резольвазой ResT [49, 50].
Состав генома
Геномная аннотация изолята Иж-4 выявила всего 1362 гена, в том числе 31 ген транспортной РНК (тРНК), один кластер из трех генов рибосомной РНК (рРНК) (5S, 16S, 23S) и три гена некодирующей РНК (нкРНК). Из 1362 генов 1222 были аннотированы как гены, кодирующие белок. Анализ показал наличие 103 (7,5%) псевдогенов в геноме Иж-4 (табл. 3). Большинство псевдогенов были результатом сдвига рамки. Количество псевдогенов различалось между геномными элементами и колебалось от 0 до 24. Наибольшее количество псевдогенов присутствовало в двух плазмидах, lp70 и lp64, и в хромосоме с 24, 23 и 22 псевдогенами соответственно.
Полная таблица
Функциональная классификация белков путем сравнения с ранее определенными кластерами ортологичных групп (COG) показала, что примерно 81% хромосомных белков и только 16% плазмидных белки Иж-4 могут быть отнесены к 25 различным категориям COG (RPS-BLAST, порог E — значение 0,01). Это подтверждает, что хромосома хорошо консервативна. Действительно, сравнение по КОГ хромосом российских изолятов с ранее секвенированными геномами американских (CT13–2396) и азиатский (FR64b) генотипы также не выявили достоверных различий.
Высокий процент белков с классификацией COG, локализованных на некоторых плазмидах, указывает на то, что некоторые плазмиды несут жизненно важные гены, которые, вероятно, кодируют белки, участвующие в основных метаболических процессах. Например, согласно нашему анализу, плазмида lp41 (41 kb) кодирует 12 белков, классифицированных как COG, а три плазмиды lp72, lp70 и lp64 кодируют 15, 10 и 9 таких белков соответственно (таблица 3). Следует отметить, что lp41 является основной плазмидой вирулентности, несущей и экспрессирующей «главные вариабельные поверхностные белки» (variable major protein, Vmps) [28].
Borrelia miyamotoi хромосома Попарное сравнение последовательностей линейной хромосомы Иж-4 с ранее секвенированными геномами FR64b (Япония), CT14D4, LB2001 и CT13-2396 (США) B. miyamotoi выявило, что средняя идентичность нуклеотидов (АНИ) между хромосомами Иж-4 и FR64b составила 99,97%, а с изолятами из США — 97,77%. Полногеномное выравнивание этих хромосом не выявило заметных геномных перестроек в виде длинных вставок\делеций, дупликаций участков и транслокаций, что подтверждает консервативный характер B. miyamotoi линейная хромосома. Однако небольшие различия были обнаружены в полиморфизмах тандемных повторов (VNTR), однонуклеотидных полиморфизмах (SNP) и небольших вставках (дополнительный файл 3: рисунки S30–S31 и таблица S2). Общее количество выявленных различий между хромосомами, что неудивительно, было разным между изолятами из разных географических регионов: Иж-4 и изоляты из США показали в среднем 18 563 различия; Иж-4 и японский изолят имели всего 122. Большинство отличий заключались в замене оснований. Мы также определили пять сайтов, содержащих VNTR (дополнительный файл 3: рисунок S30). Такие различия могут быть полезны для разработки будущих схем подтипирования для B. miyamotoi клинических изолятов.
Плазмидное типирование с помощью анализа генов паралогичных семейств (ПФ)
Идентифицированные 14 плазмидных контигов и хромосома Иж-4 были подвергнуты анализу для определения типа белков раздела и принятия решения о возможных названиях для конкретных плазмид. Для идентификации генов, гомологичных белкам репликации/поддержания плазмиды PF 32, 49, 50, 62 и 57 [51, 52], извлекали нуклеотидные последовательности открытых рамок считывания (ОРС), в том числе гены, аннотированные как псевдогены, из иж- 4 генома, а также эталонные геномы различных вида Borrelia были представлены на межпроскановую аннотацию и использованы для сравнительного филогенетического анализа (более подробное описание см. в разделе «Методы»).
Мы определили, что у Иж-4 были контиги, характеризующиеся разными генами PF (рис. 2). Методом, описанным ранее для B. burgdorferi [51], мы определили типы плазмид у Иж-4, исследуя филогенетическое родство генов PF с эталонными геномами. Гены PF 32, 49, 50, 57/62, обнаруженные на хромосоме, и несколько плазмид (lp72, lp41, lp23, lp6) были филогенетически близкородственными и образовывали монофилетические клады с генами PF, соответствующими плазмидам генома CT13–239.6 ( Дополнительный файл 4: рисунки S37–S40 ) . Несмотря на то, что в Иж-4 плазмида длиной 27 т.п.н. имела те же гены PF, что и плазмида lp23 в CT13–2396, мы выбираем для этих плазмид то же название, которое соответствует типированию плазмид в B. burgdorferi . сл [51]. Примечательно, что гены PF у Иж-4 и FR64b кластеризуются вместе в большем количестве случаев, чем у CT13–2396, что указывает на более близкое генетическое/геномное родство русского и японского B. miyamotoi изолятов, чем российских и североамериканских изолятов (включая плазмидный состав).
Схематическое изображение сегментированного генома Иж-4 с идентифицированными генами PF 32, 49, 50, 57/62. Отображаются порядок и относительное положение этих генов на плазмидах
Изображение полного размера
Мы обнаружили две плазмиды — lp70 и lp64 — которые ранее не были описаны у Borrelia . Каждая из этих плазмид несла несколько наборов генов PF, что позволяет предположить, что они образовались в результате слияния различных типов плазмид в прошлом. Плазмида lp70 Иж-4 несла две копии PF32, которые филогенетически группировались с контигами плазмиды FR64b. Однако одна из копий показала высокое сходство с PF32 плазмиды cp2 CT13–239.6 ( Дополнительный файл 4: рисунок S37 ) . Плазмида lp64 несла три набора PF 32, 49, 50, 57/62. Из них один кластер был представлен только PF50, в то время как PF57/62 был псевдогеном, а PF32 и PF49 отсутствовали. Два других набора генов содержали четыре PF гена, но один набор характеризовался наличием псевдогенов, связанных с PF 32 и 49 (рис. 2). В геноме FR64b были обнаружены две копии PF32 гена lp64, объединенные в разные филогенетические группы. Одна из копий lp64-PF32 наиболее похожа на PF32, расположенную на плазмиде pl42 из B. duttonii изолят Ly; другая копия (псевдоген) наиболее похожа на PF32, расположенную на плазмидах lpF27 B. hermsii HS1 и lp28–7 B. afzelii PKo ( Доп. файл 4: рис. S37 ) .
Плазмиды lp29, lp27, lp24, lp18–2 и lp13 содержали только одну копию PF57/62, но копия в плазмиде lp18–1 была псевдогеном PF57/62. Это согласуется с данными ранее секвенированных геномов [11]. Например, Б. Миямотои Плазмиды CT13–2396 lp30, lp20–1, lp20–2 и lp19 содержат только ген PF57/62, а плазмида cp4 несет только ген PF50 ( Дополнительный файл 4: рис. S39, S40 ) . Хотя классификация типов совместимости плазмид базировалась в основном на филогении локуса PF32, в случаях, когда этот локус отсутствовал, для типирования плазмид мы использовали PF57/62. В филогении PF57/62 плазмиды lp29, lp27, lp24, lp18–2 и lp13 Иж-4 и других изолятов B. miyamotoi сформировали кладу, отличную от большинства других видов RF и LB, за исключением B. hermsii HS1 lpG27. Почти идентичные PF57/62 были обнаружены для двух пар плазмид Иж-4: плазмиды lp29 — lp27 и lp18–1 — lp18–2. Это может поднять вопрос, действительно ли это разные плазмиды. Однако эти пары плазмид не имели других расширенных областей сходства нуклеотидов ( Дополнительный файл 3: рисунки S33, S34) за пределами локуса PF57/62, что указывает на то, что они представляют собой две разные пары плазмид. PF57/62 плазмиды lp13 сгруппированы вместе с PF57/62 lp30 CT13–2396, и ген, расположенный на плазмидном контиге (CP004259.1) FR64b. PF57/62 Izh-4 lp24 был почти идентичен гомологичному гену, расположенному на контигах плазмиды (CP004252) FR64b. Следует отметить, что кластеризация плазмид на основе генов PF32 коррелирует с группами плазмид на основе кластеризации PF57/62, указывая на схожие эволюционные закономерности между PF32 и PF57/62. Поскольку мы не выявили варианты генов PF57/62 ранее секвенированных геномов B. miyamotoi , которые были бы достаточно близки к генам PF57/62 генома Иж-4, мы решили установить названия плазмид на основе их длина.
Анализ позволил идентифицировать только две кольцевые плазмиды, каждая из которых имела длину примерно 30 т.п.н. Процент совпадения между ними составил 79%. Набор и взаимное расположение ORF между этими плазмидами коллинеарны, за исключением вариации числа генов Mlp (у cp30-1 два гена, у cp30-2 один ген) и инверсии кластера генов PF 32, 49, 50, 57/62. Обе плазмиды характеризуются наличием генов, кодирующих большую субъединицу фаговой терминазы PBSX, сайт-специфическую интегразу, что указывает на родство с профаг-родственными плазмидами [53,54,55]. Кроме того, обе кольцевые плазмиды характеризуются наличием полного набора PF 32, 49, 50, 57/62 ген. Согласно филогении генов PF32, эти две плазмиды принадлежат к разным филогенетическим кластерам. Ген PF32 плазмиды cp30–1 был более близок к гену PF32, локализованному на плазмидах pl28 ( B. duttonii Ly) и lp28–8 ( B. afzelii PKo). В свою очередь, ген PF32 плазмиды cp30–2 был филогенетически наиболее близок к гену PF32, локализованному на плазмиде lpT28 B. hermsii HS1.
Организация плазмиды вирулентности lp41
Плазмида lp41, по-видимому, играет ключевую роль в вирулентности B. miyamotoi , экспрессируя Vmps, которые позволяют бактериям ускользать от иммунной системы хозяина во время инфекции [28]. Мы провели сравнение плазмид lp41 с помощью анализа BLASTn между Иж-4 и ранее секвенированными изолятами B. miyamotoi из США (LB-2001 и CT13-2396) и Азии (FR64b). Этот анализ выявил высокую степень сходства в относительно консервативных областях 3 и 5, граничащих с вариабельной областью генов Vmp (рис. 3). Иж-4 несет ген, кодирующий белок Vlp-δ (метка локуса: CNO09_05195) после сайта экспрессии, тогда как геномы FR64b и CT13–2396 несут Vlp-γ (BOM_1113, AXh35_04655) (рис. 4), а LB-2001 несут Vsp1 (I871_B20) (рис. 5).
Сравнение нуклеотидных последовательностей плазмиды вирулентности lp41 изолятов B. miyamotoi , происходящих из Северной Америки (LB-2001, СТ13–2396), Японии (FR64b) и России (Иж-4) . Блоки, окрашенные в красный цвет (диапазон идентичности в процентах 100–70%), указывают на сходные области между плазмидами. Стрелки указывают гены и направление ORF. Блок генов Vmp представлен генами сразу после сайта экспрессии — экспрессированными генами Vmp (светло-зеленая стрелка) и неэкспрессированными генами Vmp (оранжевые стрелки). Другие ORF показаны синими стрелками
Увеличенное изображение
Рис. 4Филогенетическое разнообразие генов Vlp в геномах Izh-4, FR64b, CT13–2396 и LB-2001. Разный цвет узлов указывает на разные подсемейства Vlp: желтый — Vlp-α, красный — Vlp-γ, синий — Vlp-δ, фиолетовый — Vlp-β. ORF, выделенные жирным шрифтом, представляют собой гены, расположенные после сайта экспрессии. Теги локуса псевдогенов отмечены постфиксом «_ps». Дерево построено на основе попарного выравнивания нуклеотидных последовательностей ОРС/псевдогенов, содержащих домены, соответствующие семейству липопротеинов_2 (PF00921) или надсемейство VlsE вариабельного поверхностного антигена (SSF74748). Гены Vlp-β использовали как внешнюю группу для укоренения дерева
Увеличенное изображение
Рис. 5 Филогенетическое разнообразие генов Vsp в геномах Izh-4, FR64b, CT13–2396 и LB-2001. ORF, выделенные жирным шрифтом, представляют собой гены, расположенные после сайта экспрессии. Теги локуса псевдогенов отмечены постфиксом «_ps». Дерево было построено на основе попарного выравнивания нуклеотидных последовательностей ОРС/псевдогенов, содержащих домены, соответствующие семейству липопротеинов_6 (PF01441) или суперсемейству белка С внешней поверхности (OspC) (SSF63515). Дерево было укоренено посередине
Изображение полного размера
На левом конце плазмиды lp41 между парами изолятов FR64b — Иж-4 и СТ13-2396 — LB-2001 обнаружены минорные вставки размером 800 п.н. (данные не представлены). При этом количество и порядок генов Vmp были уникальны для каждого из изолятов (частично показано на рис. 3 и рис. 6). Кроме того, в обоих азиатских геномах, Izh-4 и FR64b, по сравнению с CT13–239, были обнаружены однонуклеотидные вариации, а также делеция 138 п.н. в межгенной области перед сайтом экспрессии.6 и LB-2001 (дополнительный файл 3: рисунок S35). Это может быть маркером дифференцировки плазмид lp41 азиатского и американского генотипов. Важно отметить, что организация сайта экспрессии последовательности не отличалась между изолятами B. miyamotoi , нуклеотидный состав сайта связывания рибосомы (RBS), сайты «-10» и «-35» были идентичными на 100% (Дополнительные файл 3: рисунок S35, внизу), что может быть очень полезным для идентификации выраженного Vmp [28].
Количество и расположение генов Vmp в геноме Иж-4
Увеличенное изображение
Внутригенетическое разнообразие вариабельных больших белков и вариабельных малых белков
Проведен поиск всех нуклеотидных последовательностей генов и псевдогенов Иж-4 для оценки их принадлежности к семейству липопротеинов в базе данных InterPro. Всего обнаружено 39 генов, кодирующих вариабельные большие белки (Vlp), из них 9 псевдогенов, и 15 генов, кодирующих вариабельные малые белки (Vsp), в том числе 5 псевдогенов. Гены Vlp и Vsp были сгруппированы по островковому типу и в основном располагались на плазмидах lp41, lp29., лп23 и лп24. Некоторые одиночные гены Vsp были расположены на плазмидах lp64, lp18–2 и lp13 (рис. 6).
Филогенетический анализ выделенных генов Vlp и псевдогенов четырех геномов B. miyamotoi показал, что гены Vlp Иж-4 образуют хорошо поддерживаемые клады: четыре клады Vlp-δ (20 генов), Vlp-γ (13 генов) , семейства Vlp-α (пять генов) и один ген на плазмиде lp29 соответствовал Vlp-β (рис. 4). Ближайшие гомологи Vlp-β с 78% аминокислотной идентичностью были идентифицированы в геномах B. crocidurae DOU (AHH07120.1) и B. hermsii (WP_064536660.1). Примечательно, что гены Vlp-β не были описаны в геномах B. miyamotoi LB2001 [28], однако аналогичные гены присутствовали в геноме CT13–2396 (AXh35_04965) и частично секвенированном геноме FR64b (BOM_1386) (Рис. 6, нижняя пурпурная ветвь).
Филогенетический анализ выделенных генов Vsp не выявил каких-либо закономерностей кластеризации (рис. 5). Однако сравнение 14 генов Vlp и 4 генов Vsp показало, что они присутствуют в двух идентичных копиях, расположенных на плазмидах lp41 и lp23. BLAST-анализ нуклеотидных последовательностей этих плазмид показал, что правые части плазмид lp41 и lp23 идентичны, с одинаковым порядком генов Vlp и Vsp и его псевдогенов (дополнительный файл 3: рисунок S36). Попарное сравнение плазмид, содержащих кластеры этих генов, не выявило сходства, подобного обнаруженному между lp41 и lp23. Такое сходство правого конца lp41 и lp23 также было обнаружено в CT13–239.6.
Сравнение последовательностей плазмид среди
изолятов B. miyamotoi Для изучения сходства плазмид между различными изолятами B. miyamotoi мы сравнили нуклеотидные последовательности трех изолятов CT13–2396, FR64b и Иж-4 ( Дополнительный файл 2: Рисунок S15 – S29). Мы выбрали эти изоляты, так как для CT13-2396 был доступен почти полный геном и для Иж-4 — полные геномы, а для FR64b черновой геном с 50 контигами был доступен в GenBank. В этих трех геномах мы обнаружили четыре общие плазмиды с высоким сходством нуклеотидов: lp72, lp41, lp23 и lp6 (таблица 4). Плазмиды lp70, lp64, lp27 и lp13 Иж-4 присутствовали только в азиатском геноме FR64b, но отсутствовали в североамериканском изоляте CT13–239.6. Плазмиды cp30–1, cp30–2, lp29, lp24, lp18–1 и lp18–2 частично присутствовали в геноме F64b и отсутствовали в CT13–2396.
Полноразмерная таблица
Филогенетический анализ
Филогенез Borrelia spp. на основе хромосомных генов
Для понимания взаимоотношений изолята Иж-4, североамериканского и азиатского изолятов B. miyamotoi , а также других видов Borrelia мы провели филогенетический анализ вновь секвенированного генома (Иж-4 ) и генома Borrelia депонированы в GenBank (дополнительный файл 1: таблица S1). На сегодняшний день эти геномы включают завершенные хромосомы и/или несколько завершенных плазмид (lp73, lp41, lp23 и lp6). Филогенетическое дерево было реконструировано с помощью конкатенированного выравнивания нуклеотидных последовательностей 249 ядерных генов, локализованных на хромосоме (минимальный процент идентичности для BLASTp 70%), идентифицированных в процессе кластеризации белков среди всех геномов Borrelia . Этот филогенетический анализ показал, что B. miyamotoi образует монофилетическую кладу внутри группы возвратного тифа и был разделен на две линии, принадлежащие к азиатскому и американскому генотипу. Азиатская линия включает Иж-4 и FR64b из Японии (рис. 7а).
a Филогенетическое дерево видов Borrelia , основанное на последовательном выравнивании нуклеотидных последовательностей 249 ядерных генов, расположенных на хромосоме. Borrelia miyamotoi кластеров с видами возвратного тифа. b Филогенетическое дерево штаммов B miyamotoi , основанное на последовательном выравнивании нуклеотидных последовательностей 719 ядерных генов. Дерево максимального правдоподобия было построено с использованием программного обеспечения RAxML с использованием модели нуклеотидных замен с гамма-распределением вариабельных положений (GTR + Γ). Полученное дерево было укоренено в средней точке с помощью Figtree (http://tree.bio.ed.ac.uk/software/figtree/). Длинные ветви, показанные не в масштабе, обозначены штриховыми линиями. Масштабная линейка показывает коэффициенты замещения
Увеличенное изображение
Для более детального анализа, т.е. для определения внутривидовых различий между изолятами B. miyamotoi , мы провели реципрокный поиск ядерных генов по BLASTp, но теперь только внутри вида B. miyamotoi . В результате было идентифицировано 719 ортологичных генов (минимальная процентная идентичность для BLASTp 80%) (рис. 7b).
Средние расстояния SNP (при последовательном выравнивании основных генов) между изолятами из Северо-Восточной Америки (CT13–2396, CT14D4, LB-2001), Японии (FR64b) и России (Иж-4) распределились следующим образом: Северо-Восточная Америка — Россия — 13 767 SNP, Северо-Восточная Америка — Япония — 13 776 SNP и Россия — Япония — 36 SNP. Среди трех северо-восточных американских изолятов было обнаружено шесть SNP.
Обсуждение
Генетически B. miyamotoi был разделен на отдельные популяции, границы которых, по-видимому, определяются ассоциацией переносчиков [2, 8, 16, 32, 33]. Исследовать генетические различия в популяций B. miyamotoi , следует использовать сравнительную геномику, но для этого требуются высококачественные эталонные геномы [56]. Хотя существует несколько черновых геномов B. miyamotoi из Северной Америки, России и Японии [11, 14, 42, 44], завершенный геном азиатского генотипа B. miyamotoi еще не завершен. Поэтому, чтобы обеспечить основу для сравнительных геномных исследований, здесь мы собрали эталонный геном для российского изолята B. miyamotoi Иж-4 с использованием доступных в настоящее время технологий длинного и короткого чтения.
Сообщалось о сборке завершенных геномов некоторых бактерий с использованием секвенирования Nanopore в качестве единственного метода [57] или сочетания длительного считывания с технологией короткого считывания Illumina [43, 58, 59]. Первоначальные сборки, сочетающие чтения Illumina и Nanopore геномов B. miyamotoi из России, дали неудовлетворительные результаты, так как не все геномные элементы (плазмиды) были правильно собраны [44]. Подобные проблемы встречались ранее с геномами B. miyamotoi , т.е. некоторые плазмиды были неполностью собраны, даже при сочетании методов длинного (PacBio SMRT) и короткого считывания (Illumina) [11]. Поэтому мы секвенировали один из доступных российских изолятов, Иж-4, с использованием технологии PacBio SMRT (в дополнение к ONT) и сравнили контиги, полученные с использованием двух технологий длительного считывания, с результатами, полученными с помощью PFGE. Кроме того, для точности консенсусные последовательности были созданы с использованием ридов последовательностей Illumina. Результатом этой стратегии стал полный эталонный геном изолята Иж-4, состоящий из одной линейной хромосомы из 906 kb, 12 линейных плазмид размером от 6 до 72 kb и две кольцевые плазмиды размером около 30 kb.
Аннотация генома B. miyamotoi изолята Иж-4 выявила всего 1362 ORF с 37 кодирующими РНК. Было предсказано, что линейная хромосома кодирует 850 ORF, и о высоком уровне консервативности линейной хромосомы свидетельствует то, что большинство этих локусов (81%) функционально классифицируются как COG. Выравнивание и сравнение хромосомы с другими российскими изолятами (данные не показаны) и изолятами из Японии (FR64b) и Северной Америки (LB-2001, CT13–239)6, CT14D4) подтвердили консервативность основной хромосомы. В этом анализе не наблюдалось ни больших, ни второстепенных перестроек, ни вставок/делеций. Как и ожидалось, сходство хромосом было более выраженным среди российских и японских изолятов с меньшим количеством SNP и более высокими значениями ANI (ANI 99,9% по сравнению с 97,7% между североамериканскими и российскими изолятами). Значения SNP между русскими/азиатскими и североамериканскими изолятами были соответственно в 1000 и 300 раз выше, чем между азиатскими и русскими изолятами.
Фракция плазмид значительно различалась, особенно между изолятами с разных континентов. Для изолята Иж-4 идентифицировано 12 линейных и две кольцевые плазмиды. По сравнению с хромосомой процент функционально классифицированных COG генов колеблется от 22 до 5%, что предполагает гораздо больше CDS с уникальной или неизвестной функцией во фракции плазмиды. Плазмиды с самым высоким процентом псевдогенов включали lp18–1 (44%), lp64 (27%), lp70 (26%), lp18–2 (23%) и lp24 (20%). В PFGE плазмиды разных размеров наблюдались для пяти дополнительных российских изолятов (т.е. Иж-5, Иж-14, Иж-16, Екат-1 и Екат-6, данные не показаны), что позволяет предположить, что высокая консервативность наблюдалась для 9 изолятов.2394 хромосома B. miyamotoi не распространяется на плазмидную фракцию. Чтобы лучше понять эволюционные отношения плазмид, мы использовали ранее описанный метод, который фокусировался на белках репликации/поддержания плазмид [51, 52].
Поскольку плазмиды Borrelia могут значительно различаться по размеру и могут подвергаться внутривидовым перестройкам, существуют проблемы идентификации плазмид из-за сходства геномного содержимого [51, 60]. Кроме того, переход плазмид от кольцевых к линейным или наоборот делает идентификацию и классификацию плазмид внутри и между боррелий видов сложных. Важность типирования плазмид подкрепляется тем фактом, что многие гены, участвующие в вирулентности и патогенности Borrelia , переносятся плазмидами.
Плазмиды в группе спирохет Лайм-боррелиоза были типизированы и классифицированы с использованием семейства паралогичных белков, описанных как белки репликации/поддержания плазмид [52, 60, 61]. Поскольку PF32 гомологичен белку ParA других бактерий, эта группа белков получила название PF32, PF49., PF50 и PF57/62 – использовали для определения типа совместимости плазмид, хотя у Borrelia PF32 может не выполнять эту роль [52, 61]. В разных плазмидах были обнаружены один или несколько наборов генов, кодирующих эти белки, в то время как в других были идентифицированы только подмножества этих молекул. Это согласуется с данными предыдущих исследований, показавших, что для спирохет Лайм-боррелиоза или изолята B. miyamotoi CT13-2396 некоторые плазмиды несут только ген PF57/62 [11, 51, 52]. Филогенетический анализ этих семейств паралогичных генов позволил нам идентифицировать плазмиды одного и того же типа совместимости (т. е. плазмиды с генами PF из одной клады) в черновых геномах североамериканских и азиатских изолята B. miyamotoi . Возможно, неудивительно, учитывая большее генетическое сходство по хромосоме и генетическую однородность популяций B. miyamotoi у одного и того же вида вектора [22, 32], у японского изолята FR64b обнаружено больше контигов с одинаковыми типами совместимости плазмид. чем в коннектикутском изоляте CT13–2396. В последнем изоляте были идентифицированы четыре плазмиды, представляющие типы совместимости с Иж-4: lp72, lp41, lp23 и lp6. Интересно, что две из этих плазмид, lp41 и lp23, содержали гены Vlp и Vsp в обоих изолятах. Всего в СТ13–2396, 23 ORF, кодирующие белки Vlp, были идентифицированы в пяти плазмидах, в то время как в Иж-4 были обнаружены четыре плазмиды, кодирующие в общей сложности 38 Vlp (включая 9 псевдогенов).
В настоящее время проводится характеристика других российских, североамериканских и европейских изолятов B. miyamotoi с использованием технологической и биоинформационной платформы, показанной в настоящем исследовании. Это подтвердит эффективность методики и поможет уточнить эволюционную историю Borrelia и связать генетические особенности B. miyamotoi с эпидемиологическими, микробиологическими, иммунологическими и клиническими особенностями этой возникающей инфекции.
Заключение
Мы собрали высококачественный эталонный геном для российского изолята B. miyamotoi , и это потребовало использования ридов двух технологий секвенирования: длинного и короткого считывания. Это обеспечивает основу для дальнейших исследований по выявлению сходств/различий между изолятами B. miyamotoi из разных географических популяций и их связи с различиями в вирулентности и патогенности для человека.
Методы
Изоляты Borrelia miyamotoi и их культурыШтамм Иж-4 выделен из крови российского пациента с острой МПК в г. Ижевске в 2016 г. и культивирован в среде MKP-F [62]. ДНК экстрагировали (различные использованные методы см. в разделах ниже) примерно из 10 9 спирохет после третьего пассажа in vitro.
Разделение плазмидной ДНК с помощью гель-электрофореза в пульсирующем поле (PFGE)
Стандартная рабочая процедура для PulseNet PFGE для энтеробактерий [63] использовалась с изменениями (Дополнительный файл 1: Метод S1). Девять внехромосомных фрагментов размером от 13 до 73 т.п.н. вырезали из геля и растворяли в буфере для растворения агарозы (Zymoresearch), экстрагировали ДНК и подвергали секвенированию Illumina.
Секвенирование всего генома и анализ данных
Секвенирование Illumina
Тотальную ДНК экстрагировали из суспензии боррелий с использованием набора DNeasy Blood & Tissue Kit для секвенирования с использованием платформы MiSeq. Для приготовления библиотеки использовали набор NexteraXT DNA Library Kit (Illumina, США). ДНК-библиотеки секвенировали с использованием набора реагентов V2 на 500 циклов на приборе MiSeq (Illumina, США). Прочтения низкого качества и последовательности адаптеров были удалены из прочтений Illumina с помощью BBTools [64]. Сборка производилась СПАдес-3.90,0 [65].
Секвенирование MinION и анализ данных
Секвенирование MinION (Oxford Nanopore Technologies, Великобритания) было выполнено ZF-Genomics, Лейден, Нидерланды. Суммарную ДНК изолята Иж-4 выделяли с помощью препарата Qiagen Tip-100 (Qiagen, Германия). Набор Native Barcoding Kit 1D (EXP-NBD103) использовали вместе с набором для секвенирования лигирования (SQK-LSK108) для приготовления библиотеки секвенирования Nanopore из тотальной ДНК. Для секвенирования использовали проточную кювету R9.4 MinION. Базовый вызов последовательностей MinION был выполнен с использованием Albacore v1.1.0; адаптеры были сняты Поречопом [66]. Canu v1.7 использовался для коррекции, обрезки и сборки de novo длинных ридов ONT с параметрами по умолчанию и размером генома 1,6 Mb. После сборки каждый контиг проверяли на наличие длинных инвертированных повторов на концах или сквозных перекрытий с использованием точечного анализа, реализованного в FlexiDot [67]. Выявленные перекрытия были проверены вручную путем выравнивания с помощью Mafft v7.271 [68] с последующим удалением. Автоматическая циркуляризация выполнялась скриптом APC (A Perfect Circle) [69].] с ручной проверкой. Сборка проекта генома была отшлифована двумя этапами коррекции. Во-первых, мы сопоставили скорректированные чтения ONT с контигами с помощью Nanopolish [70]. Затем мы скорректировали полученную консенсусную последовательность, картируя парные чтения Illumina с помощью Pilon v1.22 [45]. После картирования прочтений консенсусные последовательности были извлечены для дальнейшего анализа.
Секвенирование PacBio и анализ данных
ДНК изолята Иж-4 была отправлена в WGS с использованием секвенирования SMRT на платформе Pacific BioScience Technology. Услуга секвенирования была предоставлена основным объектом, расположенным в Норвежском центре секвенирования (NSC) (www.sequencing.uio.no). ДНК экстрагировали из 64 × 10 9 клеток с использованием набора Maxwell® 16 и набора ДНК крови Maxwell LEV (Promega, Германия). Использовался протокол подготовки библиотеки размером 20 kb. Выбор размера конечной библиотеки осуществляли с использованием гранул 0,4x Amp. Библиотека была секвенирована на приборе Pacific Biosciences RS II с использованием химии P6-C4 с временем видеосъемки 360 мин, две клетки SMRT использовались для секвенирования из-за плохой загрузки. Сборка de novo выполнялась с использованием процесса иерархической сборки генома (HGAP v3, Pacific Biosciences, SMRT Analysis Software v2.3.0) с параметрами по умолчанию (ожидаемый размер генома 1,6 Мб, минимальное целевое покрытие 15X). Программное обеспечение RS_Resequencing.1 (версия SMRT Analysis v2.3.0) использовалось для сопоставления считываний SMRT с последовательностями, чтобы исправить контиги после очистки сборки. Контиги PacBio были отшлифованы путем картирования чтений парных концов Illumina с использованием Pilon v1.22.
Определение концевых последовательностей (теломерных последовательностей) линейных репликонов
Проведено определение концевых последовательностей (теломерных последовательностей) хромосом и линейных плазмид. Для идентификации последовательностей теломер (представленных в виде палиндромных последовательностей на правом и левом концах линейных контигов) использовали собранные de novo контиги PacBio и ONT. Каждый из линейных элементов был выровнен относительно самого себя, и был выполнен точечный анализ. Для каждого выравнивания мы определили координаты контрольных точек, определенные с помощью точечного анализа, см. (Дополнительный файл 5: рисунок S41). Мы разрезали нуклеотидную последовательность на 200–1000 п.н. выше и ниже точки разрыва и проверили наличие палиндромных последовательностей с помощью инструмента Einverted пакета Emboss [71]. Если палиндромы не были обнаружены в контиге с помощью точечного анализа, мы наносили на карту обрезанные и предварительно собранные чтения PacBio на этот контиг и использовали часть прочтений, охватывающих край левого или правого концов контига, и анализировали их в инструменте Einverted.
Биоинформатический анализ
Геномные последовательности, использованные для сравнительного анализа
Таблица со всеми изолятами, использованными в этом исследовании для плазмидного типирования, сравнительной геномики и филогении, показана в дополнительной информации (Дополнительный файл 1: Таблица S1).
Расчет идентичности нуклеотидов между
хромосомами B. miyamotoi Средняя идентичность нуклеотидов (ANI) между хромосомами B. miyamotoi была рассчитана с использованием инструмента Pyani [72] с методом BLAST для выравнивания 1020 nt фрагментов входных данных. последовательности.
Определение и визуализация сходства между геномами и плазмидами
Мы использовали Circos v0.69–5 [73] и Mummer v3.0 [74], чтобы показать сходство между разными плазмидами, контигами или разными сборками. Для сравнения контиги были выровнены все против всех, и результаты выравнивания были визуализированы с помощью инструмента Circos. Учитывались только совпадающие регионы с идентичностью более 90%.
Для выявления различий в нуклеотидных последовательностях 9239Использовали 4 хромосомы B. miyamotoi различных изолятов NucDiff [75].
Чтобы определить, могут ли существовать различия в плазмиде вирулентности lp41 различных изолятов, аннотированные последовательности плазмид lp41 изолятов B. miyamotoi визуализировали и сравнивали в программе Easyfig [76].
Аннотацию генома и обозначение типов плазмид
Аннотацию собранных контигов проводили с использованием локальной версии NCBI Prokaryotic Genome Annotation Pipeline [77]. Для определения плазмидного типа конкретных контигов мы использовали принцип, предложенный Casjens et al. [51], основанный на сравнении семейств паралогичных генов (PF) 32, 49, 50, 62 и 57. Название (номенклатура), которое мы даем конкретным контигам (плазмидам), основано на родстве генов PF, идентифицированных в этих контигах, с аналогичными генами/белками ранее обозначенных плазмид в наборе эталонных геномов разных Borrelia видов с качественной аннотацией. Мы сделали вывод о родстве плазмид, обнаруженных в нашем исследовании, по сравнению с предыдущими схемами именования PF [51] и филогенезом. Мы провели поиск в базах данных белковых последовательностей с помощью программного обеспечения InterProScan, чтобы найти специфические белки PF в аннотированных геномах, таких как база данных консервативных доменов (CDD) [78], база данных белковых семейств (Pfam) [79]. ], база данных структурной и функциональной аннотации для всех белков и геномов (SUPERFAMILY) [80]. Например, было обнаружено, что PF32 гомологичен белкам в базе данных CDD с регистрационными номерами cd02038 и cd02042, PF49 связан с PF01672 в базе данных семейств белков, PF50 относится к PF02890, а PF57/62 относится к PF02414.
Наш сравнительный анализ включал следующие этапы. (1) Мы извлекли все нуклеотидные последовательности ORF, включая ORF, которые были идентифицированы как псевдогены с использованием конвейера аннотации прокариотического генома NCBI, из нашего эталона и других секвенированных геномов и поместили их в один файл. Эталонные геномы включены B. burgdorferi B31 (GCA_000008685.2), B. afzelii PKo (GCA_000222835.1), B. duttonii Ly (GCA_000019685.1), B. hermsii HS1 (GCA_001660005.1), B miyamotoi CT13–2396 (GCA_001767415.1), B. miyamotoi FR64b (GCA_000568695.2) и частично секвенированный геном Borrelia miyamotoi LB-2001 (GCA_000445425. 4). (2) Мы кластеризовали последовательности с помощью CD-HIT на уровне 90%. (3) Репрезентативная последовательность каждого кластера была подвергнута анализу InterProScan, чтобы определить, соответствует ли она определенному семейству белков в базе данных CDD, Pfam или SUPERFAMILY. (4) Впоследствии мы извлекли все последовательности из кластеров CD-HIT, представители которых соответствовали конкретным идентификаторам конкретных PF. (5) После этого мы выполнили попарное выравнивание последовательностей и реконструкцию дерева расстояний, используя сценарий pairdist [81] с 1000 бутстрепными повторами, что позволило нам понять родство между конкретными генами PF из эталонных геномов с известными именами плазмид и вновь секвенированных элементов генома в наше исследование. После обозначения типов плазмид сборки хромосом и плазмид были отправлены в GenBank.
Функциональная классификация белков путем сравнения с ранее определенными COG
Классификацию белков секвенированного генома Иж-4 на кластеры ортологичных групп (COG) проводили с помощью Perl-скрипта (cdd2cog. pl) из коллекции bac-genomics -скрипты [82].
Идентификация и филогенетический анализ генов Vmp
В качестве независимого и дополнительного анализа in silico были извлечены все нуклеотидные последовательности ORF (CDS и псевдогены) из B. miyamotoi изолят Иж-4. Затем мы подвергли все эти последовательности анализу InterProScan с использованием службы поиска совпадений InterProScan версии 5.23–62.0 с возможностью поиска в базах данных Pfam и SUPERFAMILY. Впоследствии мы получили все совпадения с семействами PF01441, SSF63515 ( белков Vsp) или PF00921, SSF74748 ( белков Vlp ) . Наконец, попарное выравнивание нуклеотидных последовательностей и филогенетический анализ были выполнены с использованием сценария pairdist с 1000 повторами начальной загрузки. Филогенетическое дерево было визуализировано с помощью модуля Ete3 Python.
Phylogenies
Идентификацию кластера ортологичных генов и получение выравнивания основного генома хромосом или конкретных плазмид проводили с помощью Roary v1. 007002 [83]. Для межвидового сравнения среди хромосом Borrelia использовалось минимум 70% идентичности для поиска BLASTp, для внутривидового сравнения хромосом B. miyamotoi это значение было установлено на 95%. Филогенетическое дерево было выведено на основе выравнивания основного генома с использованием RAxML v8.2.9.с моделью замещения нуклеотидов GTR + Γ и 1000 бутстрепных повторов. Филогенетическое дерево было визуализировано с использованием Python v2.7.11 и модуля Ete3 Python.
Доступность данных и материалов
Наборы данных, созданные в ходе текущего исследования изолята Иж-4, доступны в архиве NCBI Sequence Read Archive (SRA) (www.ncbi.nlm.nih.gov/sra/). Необработанный PacBio читает SRR7989200 (https://www.ncbi.nlm.nih.gov/sra/?term=SRR7989200), необработанный MinION читает SRR7989235 (https://www.ncbi.nlm.nih.gov/sra/?term =СРР7989235), необработанные чтения Illumina для общей ДНК-библиотеки SRR7989238 (https://www.ncbi.nlm.nih.gov/sra/?term=SRR7989238), необработанные чтения Illumina для каждого фрагмента PFGE: N1 — SRR7989237 (https:/ /trace. ncbi.nlm.nih.gov/Traces/sra/?run=SRR7989237), N2 — SRR7989232 (https://trace.ncbi.nlm.nih.gov/Traces/sra/?run=SRR7989232), N3 — SRR7989231 (https://trace.ncbi.nlm.nih.gov/Traces/sra/?run=SRR7989231), N4 — SRR7989234 (https://trace.ncbi.nlm.nih.gov/Traces/sra/? run=SRR7989234), N5 — SRR7989233 (https://trace.ncbi.nlm.nih.gov/Traces/sra/?run=SRR7989233), N6 — SRR7989244 (https://trace.ncbi.nlm.nih.gov/Traces/sra/?run=SRR7989244), N7 — SRR7989243 (https://trace.ncbi.nlm.nih.gov/Traces /sra/?run=SRR7989243), N8 — SRR7989198 (https://trace.ncbi.nlm.nih.gov/Traces/sra/?run=SRR7989198), N9 — SRR7989199 (https://trace.ncbi.nlm .nih.gov/Traces/sra/?run=SRR7989199).
Окончательный набор хромосом и плазмид для изолята Izn-4 доступен в GenBank: хромосома — CP024390.1 (https://www.ncbi.nlm.nih.gov/nuccore/CP024390), lp72 — CP024391.1 (https://www.ncbi.nlm.nih.gov/nuccore/CP024391), lp70 — CP024392.1 (https://www.ncbi.nlm.nih.gov/nuccore/CP024392.1), lp64 — CP024401.2 (https://www.ncbi.nlm.nih. gov/nuccore/CP024401.2), lp41 — CP024393.1 (https://www.ncbi.nlm.nih.gov/nuccore/CP024393.1 ), cp30-1 — CP024395.1 (https://www.ncbi.nlm.nih.gov/nuccore/CP024395.1), cp30-2 — CP040828.1 (https://www.ncbi.nlm.nih .gov/nuccore/CP040828.1), lp29 — CP024396.1 (https://www.ncbi.nlm.nih.gov/nuccore/CP024396.1), lp23 — CP024397.1 (https://www.ncbi .nlm.nih.gov/nuccore/CP024397.1), lp27 — CP024398.1 (https://www.ncbi.nlm.nih.gov/nuccore/CP024398.1), lp24 — CP024399.2 (https://www.ncbi.nlm.nih.gov/ nuccore/CP024399.2), lp18–2 — CP024400.2 (https://www.ncbi.nlm.nih.gov/nuccore/CP024400.2), lp18–1 — CP024405.2 (https://www. ncbi.nlm.nih.gov/nuccore/CP024405.2), lp13 — CP024404.1 (https://www.ncbi.nlm.nih.gov/nuccore/CP024404.1), lp6 — CP024407.1 (https: //www.ncbi.nlm.nih.gov/nuccore/CP024407.1).
Сокращения
- АОН:
Средняя идентичность нуклеотидов
- ВЗРЫВ:
Базовый инструмент локального поиска центровки
- COG:
Кластеры ортологических групп
- КП:
Циркулярная плазмида
- Фунт:
Боррелиоз Лайма
- лп:
Линейная плазмида
- ОНТ:
Оксфордские нанопоровые технологии
- ORF:
Открытая рамка считывания
- Тф:
Семейства паралогичных генов B.
burgdorferi- Телефон:
Семейство белков в базе данных семейств белков (https://pfam.xfam.org/)
- PFGE:
Гель-электрофорез в импульсном поле
- РФ:
Возвратный тиф Borrelia
- СМРТ:
Тихоокеанская биотехнология, работающая в режиме реального времени с одной молекулой
- СНП:
Однонуклеотидные полиморфизмы
- ВП:
Вариабельные крупные белки
- Вмп:
Вариабельный основной белок
- ВНТР:
Тандемные повторы с переменным числом
- Всп:
Вариабельные малые белки
Ссылки
Фукунага М., Такахаши Ю., Цурута Ю., Мацусита О., Ральф Д., Макклелланд М. и др. Генетический и фенотипический анализ Borrelia miyamotoi sp. nov., выделенный из иксодового клеща Ixodes persulcatus, переносчика болезни Лайма в Японии. Int J Syst Bacteriol. 1995;45(4):804–10.
КАС пабмед Статья Google ученый
Катлер С., Вайсье-Тауссат М., Эстрада-Пена А., Потконьяк А., Михалка А.Д., Зеллер Х. Новая боррелия на блоке: Borrelia miyamotoi — риск для здоровья человека? Евронаблюдение. 2019;24(18):1800170.
Платонов А.Е., Каран Л.С., Колясникова Н.М., Махнева Н.А., Топоркова М.Г., Малеев В.В. Люди заражаются спирохетой возвратного тифа Borrelia miyamotoi. Россия Emerg Infect Dis. 2011;17(10):1816–23.
КАС пабмед Статья Google ученый
Пуховская Н.М., Морозова О.В., Высочина Н. П., Белозерова Н.Б., Иванов Л.И. Распространенность Borrelia burgdorferi sensu lato и Borrelia miyamotoi среди иксодовых клещей на Дальнем Востоке России. Int J Parasitol Parasites Wildl. 2019; 8: 192–202.
ПабМед ПабМед Центральный Статья Google ученый
Ивабу-Ито Ю., Базарцерен Б., Наранбаатар О., Йондонжамц Э., Фуруно К., Ли К. и др. Надзор за клещами Borrelia miyamotoi и филогенетический анализ изолятов в Монголии и Японии. Клещи Tick Borne Dis. 2017;8(6):850–7.
ПабМед Статья Google ученый
Рар В, Ливанова Н, Сабитова Ю, Иголкина Ю, Ткачев С, Тикунов А и др. Естественные гибриды Ixodes persulcatus/pavlovskyi в Сибири: встречаемость в симпатрических районах и заражение широким спектром переносчиков клещей. Клещи Tick Borne Dis. 2019;10(6):101254.
ПабМед Статья Google ученый
Хаснатинов М.А., Данчинова Г.А., Такано А., Кавабата Х., Охаси Н., Масудзава Т. Распространенность Borrelia miyamotoi у Ixodes persulcatus в г. Иркутске и на прилегающих к нему территориях, Россия. Клещи Tick Borne Dis. 2016;7(2):394–7.
ПабМед Статья Google ученый
Краузе П.Дж., Фиш Д., Нарасимхан С., Барбур АГ. Инфекция Borrelia miyamotoi в природе и у человека. Клин Микробиол Инфект. 2015;21(7):631–9.
КАС пабмед ПабМед Центральный Статья Google ученый
Дибернардо А., Кот Т., Огден Н.Х., Линдси Л.Р. Распространенность инфекции Borrelia miyamotoi и сопутствующих инфекций с другими видами Borrelia spp. у клещей Ixodes scapularis, собранных в Канаде. Векторы паразитов. 2014;7:183.
ПабМед ПабМед Центральный Статья Google ученый
Ogden NH, Margos G, Aanensen DM, Drebot MA, Feil EJ, Hanincova K, et al. Исследование генотипов Borrelia burgdorferi у клещей Ixodes scapularis, собранных во время эпиднадзора в Канаде. Appl Environ Microbiol. 2011;77(10):3244–54.
КАС пабмед ПабМед Центральный Статья Google ученый
Kingry LC, Replogle A, Batra D, Rowe LA, Sexton C, Dolan M, et al. На пути к полному геному североамериканских боррелий миямотои. Объявление генома. 2017;5(5):e01557-16.
Федорова Н., Клейнджан Дж. Э., Джеймс Д., Хуэй Л. Т., Питерс Х., Лейн Р. С. Замечательное разнообразие клещей или боррелий, связанных с млекопитающими, в столичном районе залива Сан-Франциско, Калифорния. Клещи Tick Borne Dis. 2014;5(6):951–61.
ПабМед Статья Google ученый
Padgett K, Bonilla D, Kjemtrup A, Vilcins IM, Yoshimizu MH, Hui L, et al. Крупномасштабный пространственный риск и сравнительная распространенность Borrelia miyamotoi и Borrelia burgdorferi sensu lato у Ixodes pacificus. ПЛОС Один. 2014;9(10):e110853.
ПабМед ПабМед Центральный Статья КАС Google ученый
Kingry LC, Replogle A, Dolan M, Sexton C, Padgett KA, Schriefer ME. Хромосомы и большие линейные плазмидные последовательности штамма Borrelia miyamotoi, выделенного из клещей Ixodes pacificus из Калифорнии. Объявление генома. 2017;5(37):e00960-17.
Кук В.Дж., Федорова Н., Макдональд В.П., Лейн Р.С., Барбур А.Г. Уникальный штамм Borrelia miyamotoi в клещах Ixodes pacificus, Калифорния, США. Эмердж Инфекция Дис. 2016;22(12):2205–7.
ПабМед ПабМед Центральный Статья Google ученый
Вейджмейкерс А., Стааринк П.Дж., Спронг Х., Ховиус Дж. В. Borrelia miyamotoi: широко распространенная спирохета клещевого возвратного тифа. Тенденции Паразитол. 2015;31(6):260–9.
ПабМед Статья Google ученый
Геллер Ю., Назарова Л., Катаргина О., Ярвекулг Л., Фоменко Н., Головлева И. Выявление и генетическая характеристика спирохеты возвратного тифа Borrelia miyamotoi у эстонских клещей. ПЛОС Один. 2012;7(12):e51914.
КАС пабмед ПабМед Центральный Статья Google ученый
Wagemakers A, Jahfari S, de Wever B, Spanjaard L, Starink MV, de Vries HJC, et al. Borrelia miyamotoi как переносчики и хозяева в Нидерландах. Клещи Tick Borne Dis. 2017;8(3):370–4.
ПабМед Статья Google ученый
Blazejak K, Raulf MK, Janecek E, Jordan D, Fingerle V, Strube C. Сдвиги генно-видовых инфекций Borrelia burgdorferi (s. l.) у Ixodes ricinus за 10-летний период наблюдения в городе Ганновер (Германия) ) и обнаружение специфического для Borrelia miyamotoi обратного линейного блоттинга. Векторы паразитов. 2018;11(1):304.
ПабМед ПабМед Центральный Статья КАС Google ученый
Паломар А.М., Портильо А., Сантибанес П., Сантибанес С., Отео Х.А. Borrelia miyamotoi: следует ли учитывать этот возбудитель для диагностики клещевых инфекционных заболеваний в Испании? Enferm Infec Micr Cl. 2018;36(9):568–71.
Артикул Google ученый
Venczel R, Knoke L, Pavlovic M, Dzaferovic E, Vaculova T, Silaghi C, et al. Новая дуплексная ПЦР в реальном времени позволяет одновременно обнаруживать и дифференцировать Borrelia miyamotoi и Borrelia burgdorferi sensu lato. Инфекционное заболевание. 2016;44(1):47–55.
КАС пабмед Статья Google ученый
Crowder CD, Carolan HE, Rounds MA, Honig V, Mothes B, Haag H, et al. Распространенность Borrelia miyamotoi среди иксодовых клещей в Европе и США. Эмердж Инфекция Дис. 2014;20(10):1678–82.
КАС пабмед ПабМед Центральный Статья Google ученый
Barbour AG, Bunikis J, Travinsky B, Hoen AG, Diuk-Wasser MA, Fish D, et al. Разделение ниши Borrelia burgdorferi и Borrelia miyamotoi у одного и того же вида клещей-переносчиков и млекопитающих-резервуаров. Am J Trop Med Hyg. 2009 г.;81(6):1120–31.
ПабМед Статья Google ученый
Salkeld DJ, Nieto NC, Bonilla DL, Yoshimizu MH, Padgett KA. Инфекции Borrelia miyamotoi у мелких млекопитающих, Калифорния, США. Эмердж Инфекция Дис. 2018;24(12):2356–9.
ПабМед ПабМед Центральный Статья Google ученый
ван Дайвендейк Г., Койпан С., Вейджмейкерс А., Фонвиль М., Эрсоз Дж., Оэй А. и др. Личинки Ixodes ricinus передают Borrelia afzelii и B. miyamotoi позвоночным хозяевам. Векторы паразитов. 2016;9:97.
ПабМед ПабМед Центральный Статья КАС Google ученый
Хан С., Любельчик С., Хиклинг Г.Дж., Белперрон А.А., Бокенштедт Л.К., Цао Д.И. Скорость вертикальной передачи Borrelia miyamotoi в Ixodes scapularis, собранных у белохвостых оленей. Клещи Tick Borne Dis. 2019;10(3):682–9.
ПабМед Статья ПабМед Центральный Google ученый
Гофтон А.В., Маргос Г., Фингерле В., Хепнер С., Лох С.М., Райан У. и др. Полногеномный анализ Borrelia turcica и Candidatus Borrelia tachyglossi показывает рецидивирующие лихорадочные геномы с уникальными геномными связями с боррелиями болезни Лайма. Заразить Генет Эвол. 2018;66:72–81.
КАС пабмед Статья Google ученый
Барбур АГ. Множественные и разнообразные последовательности vsp и vlp у Borrelia miyamotoi, сильно переносимого клещами зоонозного патогена. ПЛОС Один. 2016;11(1):e0146283.
ПабМед ПабМед Центральный Статья КАС Google ученый
Вейджмейкерс А., Коутсвельд Дж., Нарасимхан С., Викель М., Депонте К., Блейлевенс Б. и др. Вариабельные основные белки как мишени для специфических антител против Borrelia miyamotoi. Дж Иммунол. 2016;196(10):4185–95.
КАС пабмед Статья Google ученый
Barbour AG, Burman N, Carter CJ, Kitten T, Bergstrom S. Гены вариабельного антигена возбудителя возвратного тифа Borrelia hermsii активируются добавлением промотора. Мол микробиол. 1991;5(2):489–93.
КАС пабмед Статья Google ученый
Раффель С.Дж., Баттисти Дж.М., Фишер Р.Дж., Шван Т.Г. Инактивация генов антигенной изменчивости спирохет возвратного тифа Borrelia hermsii снижает инфекционность у мышей и передачу клещами. PLoS Патог. 2014;10(4):e1004056.
ПабМед ПабМед Центральный Статья Google ученый
Буникис Дж., Цао Дж., Гарпмо Ю., Берглунд Дж., Фиш Д., Барбур АГ. Типирование штаммов группы возвратного тифа боррелий. Эмердж Инфекция Дис. 2004; 10 (9): 1661–4.
ПабМед ПабМед Центральный Статья Google ученый
Барбур АГ. Филогения возвратного тифа рода Borrelia, переносимого твердым клещом Ixodes scapularis. Заразить Генет Эвол. 2014;27:551–58.
ПабМед ПабМед Центральный Статья Google ученый
Сарксян Д.С., Платонов А.Е., Каран Л.С., Малинин И.Е., Халитова Л.И., Шахов В.И., и соавт. Клиническая картина «нового» клещевого боррелиоза, вызванного Borrelia miyamotoi. Тер Арх. 2012;84(11):34–41.
КАС пабмед Google ученый
Платонов А.Е., Топоркова М.Г., Колясникова Н.М., Стуколова О.А., Долгова А.С., Бродовикова А.В., и соавт. Клиническая картина иксодового клещевого боррелиоза, вызванного Borrelia miyamotoi, на фоне иммунного ответа на возбудителя. Тер Арх. 2017;89(11):35–43 (на рус.).
КАС пабмед Статья Google ученый
Hovius JW, de Wever B, Sohne M, Brouwer MC, Coumou J, Wagemakers A, et al. Случай менингоэнцефалита спирохетой возвратного тифа Borrelia miyamotoi в Европе. Ланцет. 2013;382(9892):658.
ПабМед ПабМед Центральный Статья Google ученый
Гульотта Дж.Л., Гётерт Х.К., Берарди В.П., Телфорд С.Р. 3-й. Менингоэнцефалит, вызванный Borrelia miyamotoi, у пациента с ослабленным иммунитетом. N Engl J Med. 2013;368(3):240–5.
КАС пабмед ПабМед Центральный Статья Google ученый
Molloy PJ, Telford SR 3rd, Chowdri HR, Lepore TJ, Gugliotta JL, Weeks KE, et al. Болезнь Borrelia miyamotoi на северо-востоке США: серия случаев. Энн Интерн Мед. 2015;163(2):91–8.
ПабМед Статья Google ученый
Боден К., Лобенштейн С., Герман Б., Маргос Г., Фингерле В. Нейроборрелиоз, связанный с Borrelia miyamotoi, у лиц с ослабленным иммунитетом. Эмердж Инфекция Дис. 2016;22(9): 1617–20.
ПабМед ПабМед Центральный Статья Google ученый
Henningsson AJ, Asgeirsson H, Hammas B, Karlsson E, Parke A, Hoornstra D, et al. Два случая менингита Borrelia miyamotoi, Швеция, 2018 г. Emerg Infect Dis. 2019; 25(10):1965–8.
ПабМед ПабМед Центральный Статья Google ученый
Хорнстра Д., Коутсвельд Дж., Спронг Х., Платонов А.Е., Ховиус Дж.В. Болезнь Borrelia miyamotoi у иммунокомпетентного пациента, Западная Европа. Эмердж Инфекция Дис. 2018;24(9): 1770–2.
ПабМед ПабМед Центральный Статья Google ученый
Хюэ Ф., Гальянчи Лангероуди А., Барбур АГ. Хромосомная последовательность Borrelia miyamotoi, некультивируемого переносимого клещами возбудителя инфекции человека. Объявление генома. 2013;1(5):e00713-13.
Маргос Г., Хепнер С., Манг С., Марошевич Д., Рейнольдс С.Е., Кребс С. и др. Затерянные в плазмидах: секвенирование следующего поколения и сложный геном клещевого возбудителя Borrelia burgdorferi. Геномика BMC. 2017;18(1):422.
КАС пабмед ПабМед Центральный Статья Google ученый
Кулешов К.В., Котсвельд Ю., Гоптарь И.А., Маркелов М.Л., Колясникова Н.М., Сарксян Д.С., и соавт. Полногеномное секвенирование шести клинических штаммов Borrelia miyamotoi, выделенных в России. Объявление генома. 2018;6(1):e01424-17.
Уокер Б.Дж., Абил Т., Ши Т., Прист М., Абуэльель А., Шактикумар С. и др. Pilon: интегрированный инструмент для комплексного обнаружения микробных вариантов и улучшения сборки генома. ПЛОС Один. 2014;9(11):e112963.
ПабМед ПабМед Центральный Статья КАС Google ученый
Casjens S, Murphy M, DeLange M, Sampson L, van Vugt R, Huang WM. Теломеры линейных хромосом спирохет болезни Лайма: последовательность нуклеотидов и возможный обмен с линейными плазмидными теломерами. Мол микробиол. 1997;26(3):581–96.
КАС пабмед Статья Google ученый
Huang WM, Robertson M, Aron J, Casjens S. Обмен теломерами между линейными репликонами Borrelia burgdorferi. J Бактериол. 2004;186(13):4134–41.
КАС пабмед ПабМед Центральный Статья Google ученый
Tourand Y, Kobryn K, Chaconas G. Специфическое для последовательности распознавание, но зависимое от положения расщепление двух различных теломер резолвазой теломер Borrelia burgdorferi, ResT. Мол микробиол. 2003;48(4):901–11.
КАС пабмед Статья Google ученый
Tourand Y, Deneke J, Moriarty TJ, Chaconas G. Характеристика и свойства реакции in vitro 19 уникальных теломер шпилек из линейных плазмид спирохеты болезни Лайма. Дж. Биол. Хим. 2009;284(11):7264–72.
КАС пабмед ПабМед Центральный Статья Google ученый
Moriarty TJ, Chaconas G. Идентификация детерминанты, обеспечивающей разрешающее использование субстрата в теломерной ресольвазе, ResT. Дж. Биол. Хим. 2009 г.;284(35):23293–301.
КАС пабмед ПабМед Центральный Статья Google ученый
Casjens SR, Mongodin EF, Qiu WG, Luft BJ, Schutzer SE, Gilcrease EB и др. Стабильность генома спирохет болезни Лайма: сравнительная геномика плазмид Borrelia burgdorferi. ПЛОС Один. 2012;7(3):e33280.
КАС пабмед ПабМед Центральный Статья Google ученый
Чаконас Г., Кобрин К. Структура, функция и эволюция линейных репликонов у боррелий. Анну Рев Микробиол. 2010;64:185–202.
КАС пабмед Статья Google ученый
Стивенсон Б., Цукерт В.Р., Акинс Д.Р. Повторение, сохранение и изменение: множественные плазмиды cp32 видов Borrelia. J Mol Microbiol Biotechnol. 2000;2(4):411–22.
КАС пабмед Google ученый
Casjens S, van Vugt R, Tilly K, Rosa PA, Stevenson B. Гомология во множестве кольцевых плазмид размером 32 килобазы, присутствующих в спирохетах болезни Лайма. J Бактериол. 1997;179(1):217–27.
КАС пабмед ПабМед Центральный Статья Google ученый
Эггерс Ч., Сэмюэлс Д.С. Молекулярные доказательства нового бактериофага Borrelia burgdorferi. J Бактериол. 1999;181(23):7308–13.
КАС пабмед ПабМед Центральный Статья Google ученый
Fraser CM, Eisen JA, Nelson KE, Paulsen IT, Salzberg SL. Ценность полного секвенирования микробного генома (вы получаете то, за что платите). J Бактериол. 2002;184(23):6403–5.
КАС пабмед ПабМед Центральный Статья Google ученый
Ломан Н.Дж., Квик Дж., Симпсон Дж.Т. Полный бактериальный геном собран de novo с использованием только данных секвенирования нанопор. Нат Методы. 2015;12(8):733–5.
КАС пабмед Статья Google ученый
Ринг Н., Абрахамс Дж. С., Джайн М., Олсен Х., Престон А., Бэгби С. Разрешение сложного генома Bordetella pertussis с использованием секвенирования нанопор со штрих-кодом. Микроб Геном. 2018;4(11):e000234.
Вик Р.Р., Джадд Л.М., Горри К.Л., Холт К.Е. Завершение сборки бактериального генома с помощью мультиплексного секвенирования MinION. Микроб Геном. 2017;3(10):e000132.
ПабМед ПабМед Центральный Google ученый
Casjens SR, Di L, Akther S, Mongodin EF, Luft BJ, Schutzer SE, et al. Первичное происхождение и диверсификация плазмид у бактерий возбудителя болезни Лайма. Геномика BMC. 2018;19(1):218.
ПабМед ПабМед Центральный Статья КАС Google ученый
Casjens SR, Gilcrease EB, Vujadinovic M, Mongodin EF, Luft BJ, Schutzer SE, et al. Плазмидное разнообразие и филогенетическая согласованность возбудителя болезни Лайма Borrelia burgdorferi. Геномика BMC. 2017;18(1):165.
ПабМед ПабМед Центральный Статья КАС Google ученый
Котсвельд Дж., Колясникова Н.М., Вейджмейкерс А., Топоркова М.Г., Сарксян Д.С., Оей А. и др. Разработка и оптимизация протокола культивирования in vitro позволяет выделять Borrelia miyamotoi от больных тяжелым клещевым возвратным тифом. Клин Микробиол Инфект. 2017;23(7):480–4.
КАС пабмед Статья Google ученый
Ribot EM, Fair MA, Gautom R, Cameron DN, Hunter SB, Swaminathan B, et al. Стандартизация протоколов гель-электрофореза в пульсирующем поле для субтипирования Escherichia coli O157:H7, Salmonella и Shigella для PulseNet. Патог пищевого происхождения Dis. 2006;3(1):59–67.
КАС пабмед Статья Google ученый
BBTools. Объединенный институт генома. https://jgi.doe.gov/data-and-tools/bbtools/. По состоянию на 9 октября 2019 г.
Банкевич А., Нурк С., Антипов Д., Гуревич А.А., Дворкин М., Куликов А.С., и др. SPAdes: новый алгоритм сборки генома и его приложения для секвенирования отдельных клеток. J Компьютерная биология. 2012;19(5):455–77.
КАС пабмед ПабМед Центральный Статья Google ученый
Porechop: адаптер-триммер для считывания Oxford Nanopore. https://github. com/rrwick/Porechop. По состоянию на 9 октября 2019 г.
FlexiDot: точечные диаграммы для анализа визуальных последовательностей. https://github.com/molbio-dresden/flexidot. По состоянию на 9 октября 2019 г.
Като К., Стэндли Д.М. Программное обеспечение MAFFT для множественного выравнивания последовательностей, версия 7: улучшения производительности и удобства использования. Мол Биол Эвол. 2013;30(4):772–80.
КАС пабмед ПабМед Центральный Статья Google ученый
APC: (a) (p) erfect (c) ircle https://github.com/jfass/apc. По состоянию на 9 октября 2019 г.
Nanopolish: алгоритмы уровня сигнала для данных MinION. https://github.com/jts/nanopolis. По состоянию на 9 октября 2019 г.
Райс П., Лонгден И., Блисби А. EMBOSS: европейский открытый пакет программного обеспечения для молекулярной биологии. Тенденции Жене. 2000;16(6):276–7.
КАС пабмед Статья Google ученый
Модуль Python для анализа идентичности средних нуклеотидов. https://github.com/widdowquinn/pyani. По состоянию на 9 октября 2019 г.
Circos: круговая визуализация. http://circos.ca/. По состоянию на 9 октября 2019 г.
Делчер А.Л., Зальцберг С.Л., Филлиппи А.М. Использование MUMmer для идентификации похожих областей в больших наборах последовательностей. Современные протоколы в биоинформатике. 2003 г.; Глава 10: Раздел 10.3.
Google ученый
Хелик К., Лагесен К., Сандве Г.К., Рогнес Т., Недербрагт А.Я. NucDiff: углубленная характеристика и аннотация различий между двумя наборами последовательностей ДНК. Биоинформатика BMC. 2017;18(1):338.
ПабМед ПабМед Центральный Статья КАС Google ученый
Салливан М.Дж., Петти Н.К., Битсон С.А. Easyfig: визуализатор сравнения геномов. Биоинформатика. 2011;27(7):1009–10.
КАС пабмед ПабМед Центральный Статья Google ученый
Конвейер аннотации генома прокариот NCBI. https://github.com/ncbi/pgap. По состоянию на 9 октября 2019 г.
Marchler-Bauer A, Derbyshire MK, Gonzales NR, Lu S, Chitsaz F, Geer LY, et al. CDD: сохраненная база данных доменов NCBI. Нуклеиновые Кислоты Res. 2015; 43 (выпуск базы данных): D222–6.
КАС пабмед Статья Google ученый
Эль-Гебали С., Мистри Дж., Бейтман А., Эдди С.Р., Лучани А., Поттер С.К. и др. База данных семейств белков Pfam в 2019 г.. Нуклеиновые Кислоты Res. 2019;47(D1):D427–D32.
КАС пабмед Статья Google ученый
Pandurangan AP, Stahlhacke J, Oates ME, Smithers B, Gough J. База данных SUPERFAMILY 2.0: значительное обновление протеома и новый веб-сервер. Нуклеиновые Кислоты Res. 2019;47(D1):D490–D4.
КАС пабмед Статья Google ученый
Парарасст. https://github.com/frederic-mahe/pairdist.git. Доступно 9Октябрь 2019 г.
Сборник скриптов для бактериальной геномики. https://github.com/aleimba/bac-genomics-scripts. По состоянию на 9 октября 2019 г.
Пейдж AJ, Cummins CA, Hunt M, Wong VK, Reuter S, Holden MT, et al. Roary: быстрый крупномасштабный пангеномный анализ прокариот. Биоинформатика. 2015;31(22):3691–3.
КАС пабмед ПабМед Центральный Статья Google ученый
Скачать ссылки
Благодарности
Служба секвенирования Pacific Biosciences была предоставлена Норвежским центром секвенирования (www. sequencing.uio.no), национальной технологической платформой, размещенной в Университете Осло и поддерживаемой программами «Функциональная геномика» и «Инфраструктура». Исследовательского совета Норвегии и Юго-восточных региональных органов здравоохранения.
Финансирование
Работа выполнена при поддержке гранта РНФ (проект № 15–15-00072-П) АЭП, КВК, НМК, ИАГ и ДСС. Немецкий национальный справочный центр по боррелиям (VF) финансировался Институтом Роберта Коха, Берлин, Германия. JWH был поддержан грантом ZonMw (номер проекта 522003007, тикает ящик Пандоры) и грантом программы ЕС Interreg для региона Северного моря в рамках проекта NorthTick.
Информация о авторе
Авторы и принадлежность
Центральный исследовательский институт эпидемиологии, Москва, 111123, Россия
Константин В. Клешов, Надежда М. Колисновая институт и Александра Э. Александра Э. Александра Э. Александра Э. Александра. Федеральный научный центр ВИЭВ», Москва, Россия
Константин В.
КулешовБаварское управление здравоохранения и безопасности пищевых продуктов, Немецкий национальный справочный центр по боррелиям, Veterinärstr. 2, 85764, Обершлайсхайм, Германия
Габриэле Маргос и Фолькер Фингерле
Медицинские центры Амстердамского университета, местонахождение Академический медицинский центр Амстердамского университета, Амстердам, Нидерланды
Йорис Котсвельд, Денис С. Сарксян и Йоппе В. Ховиус
ФНЦ им. Чумакова РАН, Москва, ул. Россия
Колясникова Надежда Михайловна
Ижевская государственная медицинская академия, Ижевск, Россия
Сарксян Денис Сергеевич
Центр стратегического планирования и управления медико-биологическими рисками для здоровья Минздрава России, Москва
Шипулин А.
В.
PubMed Google Академия- Габриэле Маргос
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Volker Fingerle
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Joris Koetsveld
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Гоптарь Ирина Анатольевна
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Михаил Л. Маркелов
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Колясникова Надежда М.
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Академия
- Сарксян Денис Сергеевич
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Кирдяшкина Нина Петровна
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Герман А. Шипулин
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
- Joppe W. Hovius
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Платонов Александр Евгеньевич
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
3 Измеровмеров 900 Исследовательский институт Минздрава труда, Москва, Россия
Гоптарь Ирина Анатольевна, Маркелов Михаил Львович, Кирдяшкина Нина Петровна
Contributions
AEP, JWH, GAS спланировали исследование. JK, AEP, NMK, DSS провели выделение и культивирование российского штамма B. miyamotoi . IAG, NPK, MLM выполнили подготовку ДНК-библиотеки и полногеномное секвенирование компанией Illumina. GM, VF, IAG предоставили PacBio полногеномное секвенирование. KVK, GM, IAG, AEP выполнили анализ PFGE, сравнительную геномику и анализ генома. GM KVK AEP JWH VF GAS написал рукопись. Все авторы читали и одобрили окончательный вариант рукописи.
Авторы переписки
Переписка с
Константин В. Кулешов или Габриэле Маргос.
Декларация этики
Утверждение этики и согласие на участие
Неприменимо.
Согласие на публикацию
Неприменимо.
Конкурирующие интересы
Авторы заявляют об отсутствии конкурирующих интересов.
Дополнительная информация
Примечание издателя
Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Дополнительная информация
Дополнительный файл 1: Таблица S1.Штаммы, использованные в этом исследовании, с регистрационными номерами NCBI Biosample.
Дополнительный файл 2: Метод S1. Разделение плазмидной ДНК с помощью гель-электрофореза в импульсном поле (PFGE). Фигурки S1 — S6. Визуальные сравнения ONT, PacBio и конечных сборок плазмид Borrelia miyamotoi изолята Иж-4. Фигурки S7 — S15. Визуальное сравнение контигов, собранных из коротких прочтений Illumina для каждого фрагмента PFGE, и окончательных сборок плазмид из Borrelia miyamotoi изолят Иж-4. Фигуры S16 – S29. Визуальное сравнение нуклеотидных последовательностей плазмид трех штаммов B. miyamotoi CT13–2396, FR64b и Иж-4 для изучения сходства плазмид.
Нуклеотидные последовательности хромосом четырех геномов B. miyamotoi (3-США, 1-Япония) были сопоставлены с хромосомой Иж-4 с помощью Mummer, а положения областей, содержащих структурные вариации, были обнаружены с помощью NucDiff и визуализированы в IGV-браузер. Рисунок S33 . Сходство участков, содержащих гены PF57/62, расположенные на плазмидах lp18–1 и lp18–2 изолята Иж-4. Рисунок S34 . Сходство участков, содержащих гены PF57/62, расположенные на плазмидах lp29 и lp27 изолята Иж-4. Рисунок S35 . Выравнивание межгенной области, расположенной выше экспрессируемого гена Vmp на lp41 FR64b, Izh-4, CT13–2396 и LB-2001. Рисунок S36. Сходство правого конца плазмид lp41 и lp23.
Дополнительный файл 4: рисунок S37.